Κορωναϊοί: Μια επισκόπηση της αναπαραγωγής και της παθογένεσής τους
Οι κοροναϊοί (CoVs), που περιβάλλουν ιούς RNA θετικής νοημοσύνης, χαρακτηρίζονται από ακίδες που μοιάζουν με κλαμπ που προεξέχουν από την επιφάνεια τους, ένα ασυνήθιστα μεγάλο γονιδίωμα RNA και μια μοναδική στρατηγική αντιγραφής.
Οι κορωναϊοί προκαλούν ποικίλες ασθένειες σε θηλαστικά και πτηνά που κυμαίνονται από εντερίτιδα σε αγελάδες και χοίρους και ασθένεια ανώτερου αναπνευστικού στα κοτόπουλα έως δυνητικά θανατηφόρες αναπνευστικές λοιμώξεις του ανθρώπου. Εδώ παρέχουμε μια σύντομη εισαγωγή σε κοροναϊούς που συζητούν την αναπαραγωγή τους και την παθογένεια τους, καθώς και τις τρέχουσες στρατηγικές πρόληψης και θεραπείας. Συζητάμε επίσης τα κρούσματα του εξαιρετικά παθογόνου κοροναϊού σοβαρού οξέος αναπνευστικού συνδρόμου (SARS-CoV) και του προσφάτως αναγνωρισμένου κοροναϊού του αναπνευστικού συνδρόμου στη Μέση Ανατολή (MERS-CoV).
1 Ταξινόμηση 2 Γονιδιωματική οργάνωσηΑνοίξτε την εικόνα σε νέο παράθυρο3 Δομή Virion 4 Coronavirus Κύκλου Ζωής 4.1 Προσάρτηση και είσοδος 4.2 Έκφραση πρωτεΐνης ρεπλικάσης 4.3 Ανατύπωση και μεταγραφή 4.4 Συναρμολόγηση και απελευθέρωση 5 Παθογένεια 5.1 Κορωναϊοί ζώων 5.2 Ανθρώπινοι κορωναϊοί 6 Διάγνωση, θεραπεία και πρόληψη 7 Συμπέρασμα
Πληροφορίες πνευματικών δικαιωμάτων Σχετικά με αυτό το πρωτόκολλο Αναφέρετε αυτό το πρωτόκολλο ως εξής: Fehr AR, Perlman S. (2015) Κορωναϊοί: Μια επισκόπηση της αντιγραφής και της παθογένεσης τους. Στο: Maier Η., Bickerton Ε., Britton Ρ. (Eds) Coronaviruses. Methods ΐη Molecular Biology, τόμος 1282. Humana Press, New York, ΝΥ
Οι κοροναϊοί (CoVs) είναι η μεγαλύτερη ομάδα ιών που ανήκουν στη σειρά Nidovirales , η οποία περιλαμβάνει τις οικογένειες Coronaviridae , Arteriviridae , Mesoniviridae και Roniviridae . Τα Coronavirinae περιλαμβάνουν μία από τις δύο υπο-οικογένειες στην οικογένεια Coronaviridae , ενώ η άλλη είναι οι Torovirinae .
Τα Coronavirinae υποδιαιρούνται περαιτέρω σε τέσσερα γένη, τους άλφα, βήτα, γάμα και δέλτα coronaviruses. Οι ιοί ταξινομήθηκαν αρχικά σε αυτά τα γένη με βάση την ορολογία, αλλά τώρα διαιρούνται με φυλογενετική συσσωμάτωση.
Όλοι οι ιοί στη σειρά Nidovirales είναι επικαλυμμένοι , μη κατακερματισμένοι ιοί RNA θετικής λογικής. Όλα περιέχουν πολύ μεγάλα γονιδιώματα για ιούς RNA, με ορισμένους ιούς που έχουν τα μεγαλύτερα αναγνωρισμένα γονιδιώματα RNA, που περιέχουν έως και 33,5 kilobase (kb) γονιδιώματα.
Άλλα κοινά χαρακτηριστικά στη σειρά των Nidovirales περιλαμβάνουν: (1) μια εξαιρετικά διατηρημένη γονιδιωματική οργάνωση, με ένα μεγάλο γονίδιο ρεπλικάσης που προηγείται των δομικών και βοηθητικών γονιδίων, (2) έκφραση πολλών μη-δομικών γονιδίων με ριβοσωμική μετατόπιση των πλαισίων. (3) αρκετές μοναδικές ή ασυνήθιστες ενζυματικές δραστικότητες που κωδικοποιούνται εντός της πολυπρωτεϊνης μεγάλης ρεπλικάσης-μεταγραφάσης. και (4) έκφραση γονιδίων κατάντη με σύνθεση 3 'nested υπο-γονιδιωματικών mRNAs. Στην πραγματικότητα, οι Nidoviralesτο όνομα της παραγγελίας προέρχεται από αυτά τα 3 'mRNAs, καθώς το nido είναι λατινικό για "φωλιά". Οι κύριες διαφορές μεταξύ των οικογενειών νιδοϊού είναι στον αριθμό, τον τύπο και τα μεγέθη των δομικών πρωτεϊνών. Αυτές οι διαφορές προκαλούν σημαντικές αλλοιώσεις στη δομή και τη μορφολογία των νουκλεοκαψιδών και των βιριόντων.
Οι κορωναϊοί περιέχουν ένα μη τμηματοποιημένο γονιδίωμα RNA θετικής λογικής των ~ 30 kb. Το γονιδίωμα περιέχει δομή καλύμματος 5 'μαζί με 3' πολυ (Α) ουρά, επιτρέποντάς του να δράσει ως mRNA για μετάφραση των πολυπρωτεϊνών ρεπλικάσης. Το γονίδιο ρεπλικάσης που κωδικοποιεί τις μη δομικές πρωτεΐνες (nsps) καταλαμβάνει τα δύο τρίτα του γονιδιώματος, περίπου 20 kb, σε αντίθεση με τις δομικές και βοηθητικές πρωτεΐνες, οι οποίες αποτελούν μόνο περίπου 10 kb του ιικού γονιδιώματος. Το 5 'άκρο του γονιδιώματος περιέχει μια αλληλουχία-οδηγό και μη μεταφρασμένη περιοχή (UTR) που περιέχει πολλαπλές δομές βρόγχου βλαστού που απαιτούνται για αναδιπλασιασμό και μεταγραφή RNA. Επιπρόσθετα, στην αρχή κάθε δομικού ή βοηθητικού γονιδίου είναι μεταγραφικές ρυθμιστικές αλληλουχίες (TRSs) που απαιτούνται για την έκφραση καθενός από αυτά τα γονίδια ( βλ. Υπότιτλος 4.3σε αναδιπλασιασμό RNA). Το 3 'UTR περιέχει επίσης δομές RNA που απαιτούνται για την αντιγραφή και σύνθεση του ιικού RNA. Η οργάνωση του γονιδιώματος του κορωναϊού είναι 5'-οδηγός-UTR-ρεπλικάση-δ (Spike) -E (φακέλου) -Μ (μεμβράνη) -Ν (νουκλεοκαψίδιο) -3 'UTR-poly (Α) ουρά με βοηθητικά γονίδια τα δομικά γονίδια στο 3 'άκρο του γονιδιώματος (Σχήμα 1 ). Οι βοηθητικές πρωτεΐνες είναι σχεδόν αποκλειστικά μη απαραίτητες για την αντιγραφή στην καλλιέργεια ιστών. Ωστόσο, μερικοί έχουν αποδειχθεί ότι έχουν σημαντικούς ρόλους στην ιική παθογένεση [ 1 ].
Εικ. 1
Γονιδιωματική οργάνωση αντιπροσωπευτικών α, β και γ CoVs. Μια απεικόνιση του γονιδιώματος MHV απεικονίζεται στην κορυφή . Οι εκτεταμένες περιοχές παρακάτω δείχνουν τις δομικές και βοηθητικές πρωτεΐνες στις 3 'περιοχές των HCoV-229E, MHV, SARS-CoV, MERS-CoV και ΙΒν. Το μέγεθος του γονιδιώματος και των επιμέρους γονιδίων προσεγγίζονται με τη χρήση του λεκτικού σήματος στο πάνω μέρος του διαγράμματος αλλά δεν έχουν σχεδιαστεί σε κλίμακα. HCoV-229E ανθρώπινο κοροναϊό 229Ε, ιό της ηπατίτιδας ποντικού MHV , κοροναϊό σοβαρής οξείας αναπνευστικής λειτουργίας SARS-CoV , κοροναϊό του αναπνευστικού συνδρόμου MERS-CoV της Μέσης Ανατολής, ιός μολυσματικής βρογχίτιδας ΙΒν
Τα βιριόνια του κοροναϊού είναι σφαιρικά με διάμετρο περίπου 125 nm όπως απεικονίζεται σε πρόσφατες μελέτες με κρυοηλεκτρονική τομογραφία και κρυο-ηλεκτρονική μικροσκοπία [ 2 , 3 ]. Το πιο χαρακτηριστικό χαρακτηριστικό των κοροναϊών είναι οι προβολές ακίδων σχήματος ράβδου που εκπέμπονται από την επιφάνεια του βιριόντος. Αυτές οι αιχμές είναι ένα καθοριστικό χαρακτηριστικό του βιριόντος και τους δίνουν την εμφάνιση ενός ηλιακού κορώνα, που προκάλεσε το όνομα, τους κοροναϊούς. Εντός του περιβλήματος του βιριόντος είναι το νουκλεοκαψίδιο. Οι κορωναϊοί έχουν ελικοειδώς συμμετρικά νουκλεοκαψίδια, τα οποία είναι ασυνήθιστα μεταξύ των ιών RNA θετικής νοημοσύνης, αλλά πολύ πιο κοινά για ιούς RNA αρνητικής λογικής.
Τα σωματίδια του κοροναϊού περιέχουν τέσσερις κύριες δομικές πρωτεΐνες. Αυτές είναι οι πρωτεΐνες ακίδας (S), μεμβράνης (Μ), περιβλήματος (Ε) και νουκλεοκαψιδίων (Ν), όλες οι οποίες κωδικοποιούνται εντός του 3 'άκρου του ιικού γονιδιώματος. Η πρωτεΐνη S (~ 150 kDa), χρησιμοποιεί μια Ν-τερματική αλληλουχία σήματος για να αποκτήσει πρόσβαση στο ER, και είναι γλυκοζυλιωμένη βαριά Ν-συνδεδεμένη. Ομοτριμερή του ιού που κωδικοποιείται πρωτεΐνη S συνθέτουν τη διακριτή δομή της ακίδας στην επιφάνεια του ιού [ 4 , 5 ]. Η τριμερής γλυκοπρωτεΐνη S είναι πρωτεΐνη σύντηξης κατηγορίας Ι [ 6 ] και μεσολαβεί στην προσκόλληση στον υποδοχέα του ξενιστή [ 7 ]. Στους περισσότερους, τους κοροναϊούς, το S διασπάται από μια πρωτεάση τύπου φουρίνης κυττάρου ξενιστή σε δύο ξεχωριστά πολυπεπτίδια σημειωμένα S1 και S2 [ 8 , 9]. Το S1 αποτελεί το μεγάλο πεδίο δέσμευσης υποδοχέα της πρωτεΐνης S, ενώ το S2 σχηματίζει τον μίσχο του μορίου της αιχμής [ 10 ].
Η πρωτεΐνη Μ είναι η πλέον άφθονη δομική πρωτεΐνη στο βιριόνιο. Είναι μια μικρή (~ 25-30 kDa) πρωτεΐνη με τρεις διαμεμβρανικές περιοχές [ 11 ] και πιστεύεται ότι δίνει στο βιριόν το σχήμα του. Έχει μια μικρή Ν-τερματική γλυκοζυλιωμένη εξωδομένη και πολύ μεγαλύτερη C-τελική ενδοδονία που εκτείνεται 6-8 nm στο ιικό σωματίδιο [ 12 ]. Παρά το γεγονός ότι έχουν εισαχθεί συν-μεταφραστικά στη μεμβράνη ER, οι περισσότερες πρωτεΐνες Μ δεν περιέχουν αλληλουχία σήματος. Πρόσφατες μελέτες υποδεικνύουν ότι η πρωτεΐνη Μ υπάρχει ως διμερές στο βιριόν και μπορεί να υιοθετήσει δύο διαφορετικές διαμόρφωση, επιτρέποντάς της να προάγει την καμπυλότητα της μεμβράνης καθώς και να δεσμεύεται με το νουκλεοκαψίδιο [ 13 ].
Η πρωτεΐνη Ε (~ 8-12 kDa) ευρίσκεται σε μικρές ποσότητες εντός του βιριόντος. Οι πρωτεΐνες του κοροναϊού Ε είναι πολύ αποκλίνουσες αλλά έχουν μια κοινή αρχιτεκτονική [ 14 ]. Η τοπολογία μεμβράνης της πρωτεΐνης Ε δεν έχει επιλυθεί πλήρως, αλλά τα περισσότερα δεδομένα υποδηλώνουν ότι είναι μια διαμεμβρανική πρωτεΐνη. Η πρωτεΐνη Ε έχει μία Ν-τερματική εξωδομένη και μία Ο-τερματική ενδοδονία και έχει δραστικότητα διαύλου ιόντων. Σε αντίθεση με άλλες δομικές πρωτεΐνες, οι ανασυνδυασμένοι ιοί που δεν έχουν την πρωτεΐνη Ε δεν είναι πάντα θανατηφόροι, αν και αυτό εξαρτάται από τον τύπο του ιού [ 15 ]. Η πρωτεΐνη Ε διευκολύνει τη συναρμολόγηση και την απελευθέρωση του ιού ( βλ. Διάκριση 4.4), αλλά έχει και άλλες λειτουργίες. Για παράδειγμα, η δραστηριότητα του διαύλου ιόντων στην πρωτεΐνη SARS-CoV E δεν απαιτείται για την αντιγραφή του ιού, αλλά απαιτείται για παθογένεση [ 16 ].
Η πρωτεΐνη Ν αποτελεί τη μόνη πρωτεΐνη που υπάρχει στο νουκλεοκαψίδιο. Αποτελείται από δύο χωριστούς τομείς, έναν Ν-τερματικό τομέα (NTD) και έναν C-τερματικό τομέα (CTD), αμφότερους ικανό να δεσμεύει RNA ίη vitro, αλλά κάθε περιοχή χρησιμοποιεί διαφορετικούς μηχανισμούς για να συνδέσει RNA. Έχει προταθεί ότι η βέλτιστη δέσμευση RNA απαιτεί συνεισφορές και από τις δύο περιοχές [ 17 , 18 ]. Η πρωτεΐνη Ν είναι επίσης έντονα φωσφορυλιωμένη [ 19 ] και έχει προταθεί φωσφορυλίωση για να προκαλέσει μια δομική αλλαγή που ενισχύει τη συγγένεια για ιικό έναντι μη ιογενούς RNA. Η πρωτεΐνη Ν δεσμεύει το γονιδίωμα του ιού σε μία διαμόρφωση τύπου σφαιριδίων σε σειρά. Δύο ειδικά υποστρώματα RNA έχουν ταυτοποιηθεί για Ν πρωτεΐνη. τα σήματα TRS [ 20 ] και το γονιδιωματικό σήμα συσκευασίας [ 21]. Το γονιδιωματικό σήμα συσκευασίας έχει βρεθεί ότι δεσμεύεται ειδικά με το δεύτερο ή C-τελικό πεδίο σύνδεσης RNA [ 22 ]. Η πρωτεΐνη Ν δεσμεύει επίσης το nsp3 [ 18 , 23 ], ένα βασικό συστατικό του συμπλόκου της ρεπλικάσης και την πρωτεΐνη Μ [ 24 ]. Αυτές οι αλληλεπιδράσεις πρωτεΐνης πιθανόν βοηθούν να προσδεθεί το γονιδίωμα του ιού στο σύμπλοκο ρεπλικάσης-μεταγραφάσης (RTC), και στη συνέχεια να συσκευάσει το ενκαψιδιωμένο γονιδίωμα σε ιικά σωματίδια.
Μια πέμπτη δομική πρωτεΐνη, η αιμαγλουτινίνη-εστεράση (ΗΕ), υπάρχει σε ένα υποσύνολο των β-κορωναϊών. Η πρωτεΐνη δρα ως αιμοσυγκολλητίνη, δεσμεύει σιαλικά οξέα στις επιφανειακές γλυκοπρωτεΐνες και περιέχει δραστικότητα ακετυλο-εστεράσης [ 25 ]. Αυτές οι δραστηριότητες πιστεύεται ότι αυξάνουν την είσοδο κυττάρων με τη μεσολάβηση πρωτεΐνης S και την εξάπλωση του ιού μέσω του βλεννογόνου [ 26 ]. Είναι ενδιαφέρον ότι η ΗΕ ενισχύει τη νευροβλεννογονία του ιού της ηπατίτιδας των ποντικών (MHV) [ 27 ]. Ωστόσο, επιλέγεται ενάντια στην καλλιέργεια ιστών για άγνωστους λόγους [ 28 ].
Η αρχική πρόσδεση του βιριονίου στο κύτταρο ξενιστή ξεκινά από αλληλεπιδράσεις μεταξύ της πρωτεΐνης S και του υποδοχέα της. Οι θέσεις των περιοχών δέσμευσης υποδοχέα (RBD) στην περιοχή S1 μιας πρωτεΐνης S του κοροναϊού ποικίλουν ανάλογα με τον ιό, με μερικές που έχουν το RBD στο Ν-τελικό άκρο του S1 (MHV), ενώ άλλες (SARS-CoV) έχουν RBD στο Ο-τελικό άκρο του S1 [ 29 , 30]. Η αλληλεπίδραση S-πρωτεΐνης-υποδοχέα είναι ο πρωταρχικός καθοριστής για έναν κορωναϊό για να μολύνει ένα είδος ξενιστή και επίσης ρυθμίζει τον τροπισμό των ιστών του ιού. Πολλοί κοροναϊοί χρησιμοποιούν πεπτιδάσες ως τον κυτταρικό υποδοχέα τους. Δεν είναι ξεκάθαρο γιατί χρησιμοποιούνται οι πεπτιδάσες, καθώς η είσοδος γίνεται ακόμη και εν απουσία της ενζυματικής περιοχής αυτών των πρωτεϊνών. Πολλοί α-κοροναϊοί χρησιμοποιούν την αμινοπεπτιδάση Ν (APN) ως υποδοχέα τους, το SARS-CoV και το HCoV-NL63 χρησιμοποιούν ως υποδοχέα το ένζυμο μετατροπής αγγειοτενσίνης 2 (ACE2), το MHV εισέρχεται μέσω CEACAM1 και το προσφάτως αναγνωρισμένο MERS-CoV δεσμεύεται με διπεπτιδυλ- πεπτιδάση 4 (DPP4) για να αποκτήσουν είσοδο σε ανθρώπινα κύτταρα ( βλέπε Πίνακα 1 για έναν κατάλογο γνωστών υποδοχέων CoV).
APN αμινοπεπτιδάση Ν, ACE2 αγγειοτασίνης-μετατρεπτικού ενζύμου 2, mCEACAM ποντικού καρκινοεμβρυονικό αντιγόνο-συναφή μορίου προσκόλλησης 1, DPP4 διπεπτιδυλοπεπτιδάσης 4, HCoV ανθρώπινο κορωναϊό, TGEV μεταδοτικής γαστρεντερίτιδας ιό, PEDV ιό επιδημίας διάρροιας χοίρου, FIPV της λοιμώδους περιτονίτιδας αιλουροειδών ιού, CCoV κορωναϊός σκύλων , Τον ιό της ηπατίτιδας μυϊκής μάζας MHV , τον κοροναϊό βοοειδών BCoV, τον κοροναϊό του σοβαρής οξείας αναπνευστικής λειτουργίας του SARS-CoV , τον κοροναϊό του αναπνευστικού συνδρόμου MERS-CoV της Μέσης Ανατολής
Μετά τη δέσμευση του υποδοχέα, ο ιός πρέπει στη συνέχεια να αποκτήσει πρόσβαση στο κυτταρόπλασμα του κυττάρου-ξενιστή. Αυτό γενικά επιτυγχάνεται με την εξαρτώμενη από οξύ πρωτεολυτική διάσπαση της πρωτεΐνης S από μία καθεψίνη, TMPRRS2 ή άλλη πρωτεάση, ακολουθούμενη από σύντηξη των ιικών και κυτταρικών μεμβρανών. Η διάσπαση της πρωτεΐνης S συμβαίνει σε δύο θέσεις εντός του τμήματος S2 της πρωτεΐνης, με την πρώτη διάσπαση σημαντική για τον διαχωρισμό των RBD και των περιοχών σύντηξης της πρωτεΐνης S [ 31 ] και η δεύτερη για την έκθεση του πεπτιδίου σύντηξης (διάσπαση στο S2 '). Η σύντηξη συμβαίνει γενικά μέσα σε οξινισμένα ενδοσώματα, αλλά μερικοί κοροναϊοί, όπως ο MHV, μπορούν να συγχωνευθούν στη μεμβράνη του πλάσματος. Η διάσπαση στο S2 'εκθέτει ένα πεπτίδιο σύντηξης που εισάγεται στη μεμβράνη, το οποίο ακολουθείται από ένωση δύο επαναλήψεων επτάδων στο S2 σχηματίζοντας μια αντιπαράλληλη δέσμη έξι έλικες [ 6]. Ο σχηματισμός αυτού του δεσμού επιτρέπει την ανάμιξη ιικών και κυτταρικών μεμβρανών, με αποτέλεσμα τη σύντηξη και τελικά την απελευθέρωση του ιικού γονιδιώματος στο κυτταρόπλασμα.
Το επόμενο βήμα στον κύκλο ζωής του κοροναϊού είναι η μετάφραση του γονιδίου ρεπλικάσης από το γονιδιωματικό RNA του ιού. Το γονίδιο της ρεπλικάσης κωδικοποιεί δύο μεγάλα ORF, rep1a και rep1b, τα οποία εκφράζουν δύο συν-τερματικές πολυπρωτεΐνες, ρρ1α και ρρ1αβ (Σχήμα 1). Για να εκφραστούν αμφότερες οι πολυπρωτεΐνες, ο ιός χρησιμοποιεί μια ολισθηρή αλληλουχία (5'-UUUAAAC-3 ') και ένα ψευδοκώτιο RNA που προκαλούν μεταβολή των ριβοσωμικών πλαισίων από το πλαίσιο ανάγνωσης rep1a στο OR1 rep1b. Στις περισσότερες περιπτώσεις, το ριβόσωμα ξετυλίγει τη δομή του ψευδοκώτου και συνεχίζει τη μετάφραση μέχρι να συναντήσει το κωδικόνιο stop rep1a. Περιστασιακά, ο ψευδοκώνας αναστέλλει το ριβόσωμα από τη συνεχή επιμήκυνση, προκαλώντας την παύση στην ολισθηρή ακολουθία, αλλάζοντας το πλαίσιο ανάγνωσης με τη μετακίνηση ενός νουκλεοτιδίου, -1 μετατόπιση πλαισίου, προτού το ριβόσωμα είναι ικανό να λιώσει τη δομή ψευδοκώτου και να επεκτείνει τη μετάφραση στο rep1b, με αποτέλεσμα τη μετάφραση του pp1ab [ 32 , 33]. Οι μελέτες in vitro προβλέπουν ότι η συχνότητα εμφάνισης ριζοσωματικών μεταβολών των πλαισίων είναι τόσο υψηλή όσο το 25%, αλλά αυτό δεν έχει προσδιοριστεί στο πλαίσιο μόλυνσης από ιό. Δεν είναι ακριβές γιατί αυτοί οι ιοί χρησιμοποιούν μετατόπιση πλαισίου για τον έλεγχο της πρωτεϊνικής έκφρασης αλλά υποτίθεται ότι είτε ελέγχει την ακριβή αναλογία πρωτεϊνών rep1b και rep1a είτε καθυστερεί την παραγωγή προϊόντων rep1b έως ότου τα προϊόντα του rep1a έχουν δημιουργήσει ένα κατάλληλο περιβάλλον για αντιγραφή RNA [ 34 ].
Οι πολυπρωτεΐνες pp1a και pp1ab περιέχουν τα nsps 1-11 και 1-16, αντίστοιχα. Στο pp1ab, το nsp11 από το pp1a γίνεται nsp12 μετά την επέκταση του ρρΐ3 στο pp1b. Ωστόσο, οι γ-κορωναϊοί δεν περιέχουν συγκρίσιμο nsp1. Αυτές οι πολυπρωτεΐνες στη συνέχεια διασπώνται στα μεμονωμένα nsps [ 35 ]. Οι κορωναϊοί κωδικοποιούν είτε δύο είτε τρεις πρωτεάσες που διασπούν τις πολυπρωτεΐνες ρεπλικάσης. Πρόκειται για τις πρωτεάσες που μοιάζουν με παπαΐνη (PLpro), που κωδικοποιούνται εντός του nsp3, και μια πρωτεάση τύπου σερίνης, η κύρια πρωτεάση ή το Mpro, που κωδικοποιείται από nsp5. Οι περισσότεροι κορωναϊοί κωδικοποιούν δύο PLpros μέσα στο nsp3, εκτός από τους γ-κορωναϊούς, SARS-CoV και MERS-CoV, οι οποίοι εκφράζουν μόνο ένα PLpro [ 36 ]. Ο PLpros διασπά τα όρια nsp1 / 2, nsp2 / 3 και nsp3 / 4, ενώ το Mpro είναι υπεύθυνο για τα υπόλοιπα 11 συμβάντα διάσπασης.
Στη συνέχεια, πολλά από τα nsps συναρμολογούνται στο σύμπλοκο ρεπλικάσης-μεταγραφάσης (RTC) για να δημιουργήσουν ένα περιβάλλον κατάλληλο για σύνθεση RNA και τελικά είναι υπεύθυνα για τον αναδιπλασιασμό και την μεταγραφή RNA των υπο-γονιδιωματικών RNAs. Τα nsps περιέχουν επίσης και άλλες περιοχές και λειτουργίες ενζύμων, συμπεριλαμβανομένων εκείνων που είναι σημαντικές για την αντιγραφή του RNA, για παράδειγμα το nsp12 κωδικοποιεί την RNA-εξαρτώμενη περιοχή RNA πολυμεράσης (RdRp) Το nsp13 κωδικοποιεί την περιοχή ελικάσης RNA και δραστικότητα 5'-τριφωσφατάσης RNA. Το nsp14 κωδικοποιεί την εξωριβονουκλεάση (ExoN) που εμπλέκεται στην πιστότητα αντιγραφής και τη δραστικότητα της Ν7-μεθυλοτρανσφεράσης. και το nsp16 κωδικοποιεί τη δραστικότητα της 2'-Ο-μεθυλοτρανσφεράσης. Εκτός από τις λειτουργίες αναπαραγωγής, έχουν αναγνωριστεί και άλλες δραστηριότητες, όπως η παρεμπόδιση των εγγενών ανοσοαποκρίσεων (nsp1, nsp16-2'-Ο-μεθυλοτρανσφεράση, nsp3-δεουβικιτανάση) για μερικά από τα nsps, ενώ άλλες έχουν άγνωστες λειτουργίες (nsp3-ADP-ριβόζη-1 "-φωσφατάση, nsp15-endoribo-νουκλεάση (NendoU)). Για μια λίστα με τις μη δομικές πρωτεΐνες και τις προτεινόμενες λειτουργίες τους,βλ. Πίνακα 2 . Είναι ενδιαφέρον ότι οι δραστηριότητες ριβονουκλεασών nsp15-NendoU και nsp14-ExoN είναι μοναδικές για τη σειρά Nidovirales και θεωρούνται γενετικοί δείκτες για αυτούς τους ιούς [ 37 ].
Πίνακας 2
Οι λειτουργίες των μη διαρθρωτικών πρωτεϊνών του coronavirus (nsps)
Πρωτεΐνη
Λειτουργία
βιβλιογραφικές αναφορές
nsp1
Προωθεί την αποικοδόμηση κυτταρικού mRNA και αποκλείει τη μετάφραση κυττάρων ξενιστή, οδηγεί σε παρεμπόδιση της έμφυτης ανοσοαπόκρισης
Ν7 ΜΤάση και 3'-5 'εξωριβονουκλεάση, ExoN. Η Ν7 ΜΤάση προσθέτει 5 'κάλυμμα σε ιικά RNAs, η δραστηριότητα ExoN είναι σημαντική για τη διόρθωση του ιικού γονιδιώματος
Ubl ουβικιτίνης-όπως, Ac όξινο, ADRP ΑϋΡ-ριβόζη-1'-φωσφατάση, PLPro παπαΐνη-όπως πρωτεάση, ΝΑΒ δέσμευσης νουκλεϊκού οξέος, SUD SARS-μοναδική περιοχή, DMVs κυστίδια διπλά-μεμβράνης, MPro κύρια πρωτεάση, RdRp RNA-εξαρτώμενη RNA πολυμεράση, ΜΤάση μεθυλτρανσφεράση, Εχο Ν ιική exoribonuclease, Nendo U ιική ενδοριβονουκλεάσης, 2'-O-ΜΤ 2'-Ο-μεθυλοτρανσφεράσης, MDA5 που συνδέονται με μελάνωμα διαφοροποίηση πρωτεΐνη 5
Η σύνθεση του ιικού RNA ακολουθεί τη μετάφραση και τη συναρμολόγηση των συμπλεγμάτων ιικής αντιγραφής. Η σύνθεση του ιικού RNA παράγει τόσο γονιδιωματικά όσο και υπο-γονιδιωματικά RNAs. Τα υπο-γονιδιωματικά RNA χρησιμεύουν ως mRNAs για τα δομικά και βοηθητικά γονίδια τα οποία βρίσκονται στη συνέχεια των πολυπρωτεϊνών ρεπλικάσης. Όλα τα υπο-γονιδιωματικά RNA με θετικό νόημα είναι 3 'συν-τερματικά με το πλήρες μήκος ιικού γονιδιώματος και έτσι σχηματίζουν ένα σύνολο από nested RNAs, μια διακριτή ιδιότητα της σειράς Nidovirales . Και τα γονιδιωματικά και υπο-γονιδιωματικά RNA παράγονται μέσω ενδιάμεσων αρνητικών κλώνων. Αυτά τα ενδιάμεσα αρνητικών κλώνων είναι μόνο περίπου 1% ως άφθονα όπως τα αντίστοιχά τους θετικής συναίσθησης και περιέχουν αμφότερες τις πολυ-ουριδινικές και αντι-ηγέτες αλληλουχίες [ 38 ].
Πολλές αλληλουχίες που ενεργούν με cis είναι σημαντικές για την αντιγραφή των ιικών RNAs. Μέσα στο 5 'UTR του γονιδιώματος υπάρχουν επτά δομές βλαστικού βρόχου που μπορεί να εκτείνονται στο γονίδιο της ρεπλικάσης 1a [ 39 , 40 , 41 , 42 ]. Το 3 'UTR περιέχει έναν βρόγχο βλαστών, ένα ψευδοκώτιο και μια υπερμεταβλητή περιοχή [ 43 , 44 , 45 , 46 ]. Είναι ενδιαφέρον ότι ο βρόχος βλαστού και ο ψευδοκώτης στο άκρο 3 'επικαλύπτονται και έτσι δεν μπορούν να σχηματιστούν ταυτόχρονα [ 44 , 47]. Επομένως, αυτές οι διαφορετικές δομές προτείνονται για τη ρύθμιση εναλλακτικών σταδίων σύνθεσης RNA, αν και ακριβώς ποια στάδια ρυθμίζονται και ο ακριβής μηχανισμός δράσης τους είναι ακόμη άγνωστος.
Ίσως η πιο πρωτότυπη πτυχή της αντιγραφής του κοροναϊού είναι το πώς τα τμήματα του οδηγού και του σώματος TRS συγχωνεύονται κατά την παραγωγή υπο-γονιδιωματικών RNAs. Αυτό αρχικά θεωρήθηκε ότι συνέβη κατά τη διάρκεια της σύνθεσης των θετικών κλώνων, αλλά τώρα πιστεύεται ότι συμβαίνει κατά τη διάρκεια της ασυνεχούς επέκτασης του αρνητικού κλώνου RNA [ 48 ]. Το τρέχον μοντέλο προτείνει ότι η RdRp παύει σε οποιαδήποτε από τις ακολουθίες TRS του σώματος (TRS-B). μετά από αυτήν την παύση, το RdRp είτε συνεχίζει την επιμήκυνση στο επόμενο TRS είτε μεταβαίνει στην ενίσχυση της αλληλουχίας οδηγός στο 5 'άκρο του γονιδιώματος καθοδηγούμενη από τη συμπληρωματικότητα του TRS-B στον οδηγό TRS (TRS-L). Πολλά αποδεικτικά στοιχεία υποστηρίζουν επί του παρόντος αυτό το μοντέλο, συμπεριλαμβανομένης της παρουσίας αλληλουχίας κατά του οδηγού στο 3 'άκρο των υπο-γονιδιωματικών RNA αρνητικών κλώνων [ 38]. Ωστόσο, εξακολουθούν να υπάρχουν πολλά ερωτήματα για τον πλήρη ορισμό του μοντέλου. Για παράδειγμα, πώς παρακάμπτει το RdRp όλες τις αλληλουχίες TRS-B για παραγωγή πλήρους μήκους γονιδιωματικού RNA γονιδίου; Επίσης, πώς ακολουθούνται οι ακολουθίες TRS-B στο TRS-L και πόση συμπληρωματικότητα είναι απαραίτητη [ 49 ]; Οι απαντήσεις σε αυτές τις ερωτήσεις και σε άλλους θα είναι απαραίτητες για να αποκτήσουν μια πλήρη προοπτική του τρόπου με τον οποίο η αντιγραφή RNA συμβαίνει σε κοροναϊούς.
Τέλος, οι κοροναϊοί είναι επίσης γνωστοί για την ικανότητά τους να ανασυνδυάζονται χρησιμοποιώντας τόσο ομόλογο όσο και μη ομόλογο ανασυνδυασμό [ 50 , 51 ]. Η ικανότητα αυτών των ιών να ανασυνδυαστούν συνδέεται με την ικανότητα μεταγωγής του κλώνου του RdRp. Ο ανασυνδυασμός παίζει σημαντικό ρόλο στην εξέλιξη του ιού και αποτελεί τη βάση για τον στοχευόμενο ανασυνδυασμό RNA, ένα εργαλείο αντεστραμμένης γενετικής που χρησιμοποιείται για την κατασκευή ιικών ανασυνδυαστών στο 3 'άκρο του γονιδιώματος.
Μετά την αντιγραφή και την υπο-γονιδιωματική σύνθεση RNA, οι ιικές δομικές πρωτεΐνες, S, Ε και Μ μεταφράζονται και εισάγονται στο ενδοπλασματικό δίκτυο (ER). Αυτές οι πρωτεΐνες κινούνται κατά μήκος της εκκριτικής οδού στο ενδιάμεσο διαμέρισμα ενδοπλασμικού δικτυώματος-Golgi (ERGIC) [ 52 , 53 ]. Εκεί, ιικά γονιδιώματα ενκαψιδιωμένα με Ν πρωτεΐνη οφθαλμού σε μεμβράνες της ERGIC που περιέχουν ιικές δομικές πρωτεΐνες, σχηματίζοντας ώριμα βιριόνια [ 54 ].
Η πρωτεΐνη Μ κατευθύνει τις περισσότερες αλληλεπιδράσεις πρωτεΐνης-πρωτεΐνης που απαιτούνται για τη συναρμολόγηση των κοροναϊών. Ωστόσο, η πρωτεΐνη Μ δεν επαρκεί για το σχηματισμό του ιού, καθώς τα σωματίδια που ομοιάζουν με ιούς (VLPs) δεν μπορούν να σχηματιστούν με έκφραση Μ πρωτεΐνης μόνο. Όταν εκφράζεται η πρωτεΐνη Μ μαζί με την πρωτεΐνη Ε, σχηματίζονται VLPs, υποδηλώνοντας ότι αυτές οι δύο πρωτεΐνες λειτουργούν μαζί για να παράγουν φακέλους κοροναϊού [ 55 ]. Η πρωτεΐνη Ν ενισχύει τον σχηματισμό VLP, υποδηλώνοντας ότι η σύντηξη ενκαψιδιωμένων γονιδιωμάτων στο ERGIC ενισχύει την εμφάνιση ιού [ 56 ]. Η πρωτεΐνη S ενσωματώνεται σε βιριόνια σε αυτό το βήμα, αλλά δεν απαιτείται για συναρμολόγηση. Η ικανότητα της πρωτεΐνης S να κυκλοφορεί στην ERGIC και να αλληλεπιδρά με την πρωτεΐνη Μ είναι κρίσιμη για την ενσωμάτωσή της σε βιριόνια.
Ενώ η πρωτεΐνη Μ είναι σχετικά άφθονη, η πρωτεΐνη Ε είναι παρούσα μόνο σε μικρές ποσότητες στο ιοσωμάτιο. Έτσι, είναι πιθανό ότι οι αλληλεπιδράσεις της πρωτεΐνης Μ παρέχουν την ώθηση για την ωρίμανση του φακέλλου. Δεν είναι γνωστό πώς η πρωτεΐνη Ε βοηθά την πρωτεΐνη Μ στη συναρμολόγηση του βιριόντος και έχουν προταθεί διάφορες δυνατότητες. Κάποια εργασία έδειξε ρόλο για την πρωτεΐνη Ε στην πρόκληση καμπυλότητας της μεμβράνης [ 57 , 58 , 59 ], αν και άλλοι έχουν προτείνει ότι η πρωτεΐνη Ε παρεμποδίζει τη συσσωμάτωση της πρωτεΐνης Μ [ 60 ]. Η πρωτεΐνη Ε μπορεί επίσης να έχει ξεχωριστό ρόλο στην προαγωγή της απελευθέρωσης του ιού με μεταβολή της εκκριτικής οδού του ξενιστή [ 61 ].
Η πρωτεΐνη Μ προσδένεται επίσης στο νουκλεοκαψίδιο, και αυτή η αλληλεπίδραση προάγει την ολοκλήρωση της συναρμολόγησης του ιού. Αυτές οι αλληλεπιδράσεις έχουν χαρτογραφηθεί στο Ο-τελικό άκρο της ενδοδονίας του Μ με CTD της Ν-πρωτεΐνης [ 62 ]. Ωστόσο, είναι ασαφές ακριβώς πώς το νουκλεοκαψίδιο που συμπλέκεται με το RNA του ιωδιούχου κυκλοφορεί στο ERGIC για να αλληλεπιδράσει με την πρωτεΐνη Μ και να ενσωματωθεί στον ιικό φάκελο. Ένα άλλο σημαντικό ζήτημα είναι το πώς η πρωτεΐνη Ν συσκευάζει επιλεκτικά μόνο γονιδιώματα πλήρους μήκους θετικής νοημοσύνης μεταξύ των πολλών διαφορετικών ειδών RNA που παράγονται κατά τη διάρκεια της μόλυνσης. Ένα σήμα συσκευασίας για την MHV έχει ταυτοποιηθεί στην κωδικεύουσα αλληλουχία nsp15, αλλά η μετάλλαξη αυτού του σήματος δεν φαίνεται να επηρεάζει την παραγωγή ιού και δεν έχει καθοριστεί ένας μηχανισμός για τον τρόπο με τον οποίο λειτουργεί αυτό το σήμα συσκευασίας [ 22]. Επιπλέον, οι περισσότεροι κορωναϊοί δεν περιέχουν παρόμοιες αλληλουχίες σε αυτόν τον τόπο, υποδεικνύοντας ότι η συσκευασία μπορεί να είναι εξειδικευμένη στον ιό.
Μετά τη συναρμολόγηση, τα βιριόνια μεταφέρονται στην κυτταρική επιφάνεια σε κυστίδια και απελευθερώνονται με εξωκύτωση. Δεν είναι γνωστό αν τα βιριόνια χρησιμοποιούν την παραδοσιακή οδό μεταφοράς μεγάλου φορτίου από το Golgi ή αν ο ιός έχει εκτρέψει μια ξεχωριστή, μοναδική οδό για τη δική του έξοδο. Σε αρκετούς κοροναϊούς, η πρωτεΐνη S που δεν συναρμολογείται σε βιριόνια μεταβαίνει στην κυτταρική επιφάνεια όπου μεσολαβεί στη σύντηξη κυττάρου-κυττάρου μεταξύ μολυσμένων κυττάρων και γειτονικών, μη μολυσμένων κυττάρων. Αυτό οδηγεί στο σχηματισμό γιγαντιαίων πολυπυρηνικών κυττάρων, τα οποία επιτρέπουν στον ιό να εξαπλωθεί μέσα σε μολυσμένο οργανισμό χωρίς να ανιχνευθεί ή να εξουδετερωθεί από αντισώματα ειδικά για τον ιό.
Οι κοροναϊοί προκαλούν μεγάλη ποικιλία ασθενειών σε ζώα και η ικανότητά τους να προκαλούν σοβαρές ασθένειες στα ζώα και τα ζώα συντροφιάς όπως χοίροι, αγελάδες, κοτόπουλα, σκύλοι και γάτες οδήγησαν σε σημαντική έρευνα για τους ιούς αυτούς κατά το τελευταίο μισό του εικοστού αιώνα. Για παράδειγμα, ο ιός της μεταδοτικής γαστρεντερίτιδας (TGEV) και ο ιός της ριπιδαιμικής νόσου των χοίρων (PEDV) προκαλούν σοβαρή γαστρεντερίτιδα στα νεαρά χοιρίδια, οδηγώντας σε σημαντική νοσηρότητα, θνησιμότητα και τελικά οικονομικές απώλειες. Το PEDV εμφανίστηκε πρόσφατα στη Βόρεια Αμερική για πρώτη φορά, προκαλώντας σημαντικές απώλειες νεαρών χοιριδίων. Ο ιός αιμοσυγκολλητικής εγκεφαλομυελίτιδας χοίρου (PHEV) οδηγεί ως επί το πλείστον σε εντερική λοίμωξη αλλά έχει την ικανότητα να μολύνει το νευρικό σύστημα προκαλώντας εγκεφαλίτιδα, εμετό και σπατάλη σε χοίρους. Ο εντερικός κοροναϊός της γάτας (FCoV) προκαλεί ήπια ή ασυμπτωματική μόλυνση σε οικιακές γάτες, αλλά κατά τη διάρκεια επίμονης μόλυνσης, η μετάλλαξη μετασχηματίζει τον ιό σε ένα εξαιρετικά λοιμογόνο στέλεχος FCoV, ιό της μολυσματικής μολυσματικής περιτονίτιδας (FIPV), που οδηγεί στην ανάπτυξη μιας θανατηφόρου νόσου λοιμώδη περιτονίτιδα αιλουροειδών (FIP). Το FIP έχει υγρές και ξηρές μορφές, με ομοιότητες με την ασθένεια του ανθρώπου, τη σαρκοείδωση. Το FIPV είναι τροπικό μακροφάγο και πιστεύεται ότι προκαλεί ανώμαλη έκφραση κυτοκίνης και / ή χημειοκίνης και εξάντληση λεμφοκυττάρων, με αποτέλεσμα θανατηφόρο νόσημα [ με ομοιότητες με την ασθένεια του ανθρώπου, τη σαρκοείδωση. Το FIPV είναι τροπικό μακροφάγο και πιστεύεται ότι προκαλεί ανώμαλη έκφραση κυτοκίνης και / ή χημειοκίνης και εξάντληση λεμφοκυττάρων, με αποτέλεσμα θανατηφόρο νόσημα [ με ομοιότητες με την ασθένεια του ανθρώπου, τη σαρκοείδωση. Το FIPV είναι τροπικό μακροφάγο και πιστεύεται ότι προκαλεί ανώμαλη έκφραση κυτοκίνης και / ή χημειοκίνης και εξάντληση λεμφοκυττάρων, με αποτέλεσμα θανατηφόρο νόσημα [63 ]. Ωστόσο, απαιτείται περαιτέρω έρευνα για να επιβεβαιωθεί αυτή η υπόθεση. Το βόειο CoV, το Rat CoV και ο ιός της λοιμώδους βρογχίτιδας (IBV) προκαλούν ήπια έως σοβαρή λοιμώξεις της αναπνευστικής οδού σε βοοειδή, αρουραίους και κοτόπουλα, αντίστοιχα. Το βοοειδές CoV προκαλεί σημαντικές απώλειες στη βιομηχανία βοοειδών και επίσης έχει εξαπλωθεί για να μολύνει μια ποικιλία από μηρυκαστικά, συμπεριλαμβανομένων των λοφίων, των ελαφιών και των καμήλων. Εκτός από τη σοβαρή αναπνευστική ασθένεια, ο ιός προκαλεί διάρροια («χειμερινή δυσεντερία» και «ναυτικό πυρετό»), όλα οδηγούν σε απώλεια βάρους, αφυδάτωση, μειωμένη παραγωγή γάλακτος και κατάθλιψη [63]. Μερικά στελέχη του IBV, ενός γ-κορωναϊού, επηρεάζουν επίσης την ουρογεννητική οδό κοτόπουλων που προκαλεί νεφρική νόσο. Η μόλυνση του αναπαραγωγικού συστήματος με IBV μειώνει σημαντικά την παραγωγή αυγών προκαλώντας σημαντικές απώλειες στον κλάδο παραγωγής αυγών κάθε χρόνο [ 63 ]. Πιο πρόσφατα, ένας νέος κοροναϊός που ονομάζεται SW1 έχει ταυτοποιηθεί σε μια νεκρή φάλαινα Beluga [ 64]. Μεγάλος αριθμός σωματιδίων ιού εντοπίστηκαν στο ήπαρ της νεκρωμένης φάλαινας με αναπνευστική νόσο και οξεία ηπατική ανεπάρκεια. Αν και οι ηλεκτρονικές μικροσκοπικές εικόνες δεν ήταν επαρκείς για να αναγνωρίσουν τον ιό ως κορωναϊό, η αλληλούχιση του ιστού του ήπατος αναγνώρισε σαφώς τον ιό ως κορωναϊό. Στη συνέχεια προσδιορίστηκε ότι είναι γ-κορωναϊός με βάση τη φυλογενετική ανάλυση, αλλά δεν έχει ακόμη επαληθευτεί πειραματικά ότι αυτός ο ιός είναι στην πραγματικότητα αιτιολογικός παράγοντας ασθένειας σε φάλαινες. Επιπρόσθετα, υπήρξε έντονο ενδιαφέρον για την ταυτοποίηση νέων CoVs νυχτερίδας, καθώς αυτοί είναι οι πιθανότεροι πρόγονοι για τα SARS-CoV και MERS-CoV και έχουν εντοπιστεί εκατοντάδες νέοι κοροναϊοί κατά τη διάρκεια της τελευταίας δεκαετίας [ 65 ]. Τέλος, μια άλλη νέα οικογένεια νιντοϊών, Mesoniviridae, αναγνωρίστηκε πρόσφατα ως οι πρώτοι νιοϊοί για να μολύνουν αποκλειστικά ξενιστές εντόμων [ 66 , 67 ]. Αυτοί οι ιοί είναι εξαιρετικά αποκλίνοντες από άλλους νιοϊούς, αλλά είναι πολύ στενοί συγγενείς προς τους ρονινοϊούς. Στο μέγεθος, είναι ~ 20 kb, που πέφτουν μεταξύ μεγάλων και μικρών νινοϊών. Είναι ενδιαφέρον ότι αυτοί οι ιοί δεν κωδικοποιούν για μια ενδοριβονουκλεάση, η οποία υπάρχει σε όλους τους άλλους νιοϊούς. Αυτά τα χαρακτηριστικά υποδεικνύουν ότι αυτοί οι ιοί είναι το πρωτότυπο μιας νέας οικογένειας nidovirus και μπορεί να είναι ένας σύνδεσμος που λείπει κατά τη μετάβαση από μικρούς σε μεγάλους νιοϊούς.
Ο πιο βαριά μελετώμενος ζωικός κοροναϊός είναι ο ιός της ηπατίτιδας του μυοειδούς (MHV), ο οποίος προκαλεί μια ποικιλία αποτελεσμάτων σε ποντίκια, συμπεριλαμβανομένων των αναπνευστικών, εντερικών, ηπατικών και νευρολογικών λοιμώξεων. Αυτές οι μολύνσεις συχνά χρησιμεύουν ως εξαιρετικά χρήσιμα μοντέλα ασθένειας. Για παράδειγμα, το MHV-1 προκαλεί σοβαρή αναπνευστική ασθένεια σε ευαίσθητους A / J και C3H / HeJ ποντικούς, A59 και MHV-3 προκαλούν σοβαρή ηπατίτιδα, ενώ το JHMV προκαλεί σοβαρή εγκεφαλίτιδα. Είναι ενδιαφέρον ότι η MHV-3 επάγει κυτταρική βλάβη μέσω της ενεργοποίησης του καταρράκτη πήξης [ 68]. Πιο συγκεκριμένα, η A59 και οι εξασθενημένες εκδόσεις του JHMV προκαλούν μια χρόνια ασθένεια απομυελινωτής που φέρει ομοιότητες με τη σκλήρυνση κατά πλάκας (MS), καθιστώντας τη μόλυνση MHV ένα από τα καλύτερα μοντέλα για αυτή την εξασθενητική ανθρώπινη ασθένεια. Οι πρώτες μελέτες πρότειναν ότι η απομυελίνωση εξαρτάται από τον ιικό αναδιπλασιασμό στα ολιγοδενδροκύτταρα στον εγκέφαλο και στον νωτιαίο μυελό [ 69 , 70 ]. Ωστόσο, πιο πρόσφατες αναφορές καταδεικνύουν σαφώς ότι η ασθένεια προκαλείται από ανοσοποίηση. Τα ακτινοβολημένα ποντίκια ή τα ανοσοανεπαρκή (χωρίς κύτταρα Τ και Β) δεν αναπτύσσουν απομυελίνωση, αλλά η προσθήκη ειδικών για τον ιό Τ κυττάρων αποκαθιστά την ανάπτυξη απομυελίνωσης [ 71 , 72]. Επιπρόσθετα, η απομυελίνωση συνοδεύεται από μεγάλη εισροή μακροφάγων και μικρογλοιών που μπορούν να φαγοκυττάρω μολυσμένη μυελίνη [ 73 ], αν και δεν είναι γνωστό ποια είναι τα σήματα ότι τα άμεσα κύτταρα του ανοσοποιητικού συστήματος καταστρέφουν τη μυελίνη. Τέλος, η MHV μπορεί να μελετηθεί υπό εργαστηριακές συνθήκες BSL2, σε αντίθεση με τα SARS-CoV ή MERS-CoV, τα οποία απαιτούν εργαστήριο BSL3 και παρέχουν μεγάλο αριθμό κατάλληλων ζωικών μοντέλων. Αυτοί οι παράγοντες καθιστούν το MHV ιδανικό μοντέλο για τη μελέτη των βασικών αρχών της αντιγραφής του ιού σε κύτταρα ιστικής καλλιέργειας καθώς και για τη μελέτη της παθογένειας και της ανοσοαπόκρισης σε κοροναϊούς.
Πριν από την έκρηξη του SARS-CoV, οι κοροναϊοί θεωρούνταν ότι προκαλούσαν ήπιες, αυτοπεριοριζόμενες αναπνευστικές λοιμώξεις στους ανθρώπους. Δύο από αυτούς τους ανθρώπινους κοροναϊούς είναι α-κορωναϊοί, HCoV-229E και HCoV-NL63, ενώ οι άλλοι δύο είναι β-κορωναϊοί, HCoV-OC43 και HCoV-HKU1. HCoV-229E και HCoV-OC43 απομονώθηκαν πριν από περίπου 50 χρόνια [ 74 , 75 , 76 ], ενώ τα HCoV-NL63 και HCoV-HKU1 ταυτοποιήθηκαν πρόσφατα μετά την επιδημία SARS-CoV [ 77 , 78]. Αυτοί οι ιοί είναι ενδημικοί στους ανθρώπινους πληθυσμούς, προκαλώντας 15-30% των λοιμώξεων του αναπνευστικού συστήματος κάθε χρόνο. Προκαλούν πιο σοβαρή νόσο στα νεογνά, στους ηλικιωμένους και σε άτομα με υποκείμενες ασθένειες, με μεγαλύτερη συχνότητα εμφάνισης λοίμωξης του κατώτερου αναπνευστικού συστήματος σε αυτούς τους πληθυσμούς. Το HCoV-NL63 σχετίζεται επίσης με οξεία λαρυγγοτραχειίτιδα (κροπάγος) [ 79 ]. Μια ενδιαφέρουσα πτυχή αυτών των ιών είναι οι διαφορές τους όσον αφορά την ανοχή στη γενετική μεταβλητότητα. Τα απομονώματα HCoV-229E από όλο τον κόσμο έχουν μόνο ελάχιστη απόκλιση αλληλουχίας [ 80 ], ενώ τα HCoV-OC43 απομονώνονται από την ίδια θέση αλλά απομονώνονται σε διαφορετικά έτη παρουσιάζουν σημαντική γενετική μεταβλητότητα [ 81]. Αυτό πιθανόν να εξηγεί την ανικανότητα του HCoV-229E να διασχίσει το φράγμα των ειδών για να μολύνει ποντίκια ενώ το HCoV-OC43 και ο στενά συνδεδεμένος βόειος κοροναϊός, BCoV, είναι ικανοί να μολύνουν ποντίκια και διάφορα είδη μηρυκαστικών. Με βάση την ικανότητα της MHV να προκαλεί απομυελωτική ασθένεια, έχει προταθεί ότι τα ανθρώπινα CoVs μπορεί να εμπλέκονται στην ανάπτυξη σκλήρυνσης κατά πλάκας (MS). Ωστόσο, μέχρι σήμερα δεν υπάρχουν ενδείξεις ότι τα ανθρώπινα CoVs διαδραματίζουν σημαντικό ρόλο στα κράτη μέλη.
Ο SARS-CoV, ένας β-κοροναϊός της ομάδας 2b, αναγνωρίστηκε ως ο αιτιολογικός παράγοντας της επιδημίας Σοβαρής Οξείας Αναπνευστικής Συγκυρίας (SARS) που εμφανίστηκε το 2002-2003 στην επαρχία Γκουανγκντόνγκ της Κίνας. Είναι η πιο σοβαρή ανθρώπινη ασθένεια που προκαλείται από οποιονδήποτε κοροναϊό. Κατά τη διάρκεια της επιδημίας 2002-2003 περίπου 8.098 περιπτώσεις εμφανίστηκαν με 774 θανάτους, με αποτέλεσμα ποσοστό θνησιμότητας 9%. Το ποσοστό αυτό ήταν πολύ υψηλότερο στα ηλικιωμένα άτομα, με τα ποσοστά θνησιμότητας να προσεγγίζουν το 50% σε άτομα άνω των 60 ετών. Επιπλέον, το ξέσπασμα είχε ως αποτέλεσμα την απώλεια περίπου 40 δισεκατομμυρίων δολαρίων στην οικονομική δραστηριότητα, καθώς ο ιός σχεδόν έκλεισε πολλές δραστηριότητες στη Νοτιοανατολική Ασία και το Τορόντο του Καναδά για αρκετούς μήνες. Το ξέσπασμα ξεκίνησε σε ένα ξενοδοχείο στο Χονγκ Κονγκ και τελικά εξαπλώθηκε σε περισσότερες από δύο χώρες. Κατά τη διάρκεια της επιδημίας,82 ]. Εντούτοις, είναι ευρέως αποδεκτό ότι το SARS-CoV προέρχεται από νυχτερίδες, καθώς ένας μεγάλος αριθμός κινεζικών νυχτερίδων πετάλων περιέχει αλληλουχίες σχετιζόμενες με το SARS CoVs και περιέχουν ορολογικές ενδείξεις για προηγούμενη μόλυνση με σχετικό CoV [83 ,84 ]. Στην πραγματικότητα, πρόσφατα εντοπίστηκαν δύο νέοι CoVs σχετιζόμενοι με το SARS, οι οποίοι συσχετίζονται περισσότερο με το SARS-CoV από οποιονδήποτε άλλο ιό μέχρι σήμερα [85 ]. Βρέθηκαν επίσης ότι χρησιμοποιούν τον ίδιο υποδοχέα όπως ο ανθρώπινος ιός, το ένζυμο μετατροπής αγγειοτενσίνης 2 (ACE2), παρέχοντας περαιτέρω στοιχεία ότι το SARS-CoV προέρχεται από νυχτερίδες. Αν και ορισμένα άτομα σε υγρές αγορές ζώων είχαν ορολογική ένδειξη μόλυνσης με SARS-CoV πριν από την εμφάνιση της νόσου, αυτά τα άτομα δεν είχαν εμφανή συμπτώματα [82]. Έτσι, είναι πιθανό ότι ένας πολύ συγγενής ιός κυκλοφόρησε στις αγορές υγρών ζώων για αρκετά χρόνια, πριν από μια σειρά παραγόντων που διευκόλυναν την εξάπλωσή του στον μεγαλύτερο πληθυσμό.
Η μετάδοση του SARS-CoV ήταν σχετικά ανεπαρκής, καθώς απλώθηκε μόνο μέσω άμεσης επαφής με μολυσμένα άτομα μετά την εμφάνιση της ασθένειας. Έτσι, το ξέσπασμα συμπεριλήφθηκε σε μεγάλο βαθμό στα νοικοκυριά και τις εγκαταστάσεις υγειονομικής περίθαλψης [ 86 ], εκτός από μερικές περιπτώσεις υπερέκφρασης γεγονότων όπου ένα άτομο ήταν σε θέση να μολύνει πολλαπλές επαφές λόγω αυξημένης ανάπτυξης υψηλών ιικών φορτίων ή ικανότητας για αερολύμωση ιού. Ως αποτέλεσμα της σχετικά ανεπαρκούς μετάδοσης του SARS-CoV, η επιδημία ήταν ελεγχόμενη μέσω της χρήσης της απομόνωσης. Μόνο ένας μικρός αριθμός περιστατικών SARS συνέβη μετά τον έλεγχο της εστίας τον Ιούνιο του 2003.
Το SARS-CoV μολύνει κυρίως επιθηλιακά κύτταρα εντός του πνεύμονα. Ο ιός είναι ικανός να εισέλθει σε μακροφάγα και δενδριτικά κύτταρα, αλλά οδηγεί μόνο σε αποτυχημένη μόλυνση [ 87 , 88 ]. Παρά ταύτα, η μόλυνση αυτών των κυτταρικών τύπων μπορεί να είναι σημαντική στην πρόκληση προ-φλεγμονωδών κυτοκινών που μπορεί να συνεισφέρουν στην ασθένεια [ 89 ]. Στην πραγματικότητα, πολλές κυτοκίνες και χημειοκίνες παράγονται από αυτούς τους τύπους κυττάρων και είναι αυξημένες στον ορό ασθενών με μόλυνση από SARS-CoV [ 90]. Ο ακριβής μηχανισμός της πνευμονικής βλάβης και η αιτία της σοβαρής ασθένειας στους ανθρώπους παραμένει απροσδιόριστος. Οι ιικοί τίτλοι φαίνεται να μειώνονται όταν αναπτύσσεται σοβαρή ασθένεια τόσο σε ανθρώπους όσο και σε διάφορα ζωικά μοντέλα της νόσου. Επιπλέον, τα ζώα που έχουν μολυνθεί από στελέχη SARS-CoV προσαρμοσμένα σε τρωκτικά εμφανίζουν παρόμοια κλινικά χαρακτηριστικά με τη νόσο του ανθρώπου, συμπεριλαμβανομένης μιας εξαρτώμενης από την ηλικία αύξησης της σοβαρότητας της νόσου [ 91 ]. Αυτά τα ζώα παρουσιάζουν επίσης αυξημένα επίπεδα προφλεγμονωδών κυτοκινών και μειωμένες αποκρίσεις Τ-κυττάρων, γεγονός που υποδηλώνει πιθανό ανοσοπαθολογικό μηχανισμό της νόσου [ 92 , 93 ].
Ενώ η επιδημία SARS-CoV ελέγχθηκε το 2003 και ο ιός δεν επιστρέφει από τότε, ένα νέο ανθρώπινο CoV προέκυψε στη Μέση Ανατολή το 2012. Αυτός ο ιός, που ονομάστηκε Αναπνευστικό Σύνδρομο Μέσης Ανατολής-CoV (MERS-CoV), βρέθηκε να είναι ο αιτιολογικός παράγοντας σε μια σειρά από υψηλής παθογονικότητας λοιμώξεις του αναπνευστικού συστήματος στη Σαουδική Αραβία και σε άλλες χώρες της Μέσης Ανατολής [ 94]. Με βάση το υψηλό ποσοστό θνησιμότητας της τάξης του ~ 50% στα αρχικά στάδια της εκδήλωσης, φοβήθηκε ότι ο ιός θα οδηγήσει σε μια πολύ σοβαρή εστία. Ωστόσο, η επιδημία δεν επιταχύνθηκε το 2013, παρόλο που οι σποραδικές περιπτώσεις συνεχίστηκαν καθ 'όλη τη διάρκεια του χρόνου. Τον Απρίλιο του 2014, σημειώθηκε μια αιχμή πάνω από 200 περιστατικών και σχεδόν 40 θανάτους, προκαλώντας φόβους ότι ο ιός είχε μεταλλαχθεί και ήταν πιο ικανός για μετάδοση από άνθρωπο σε άνθρωπο. Πιθανότατα, ο αυξημένος αριθμός περιπτώσεων προέκυψε από βελτιωμένες μεθόδους ανίχνευσης και αναφοράς, σε συνδυασμό με την εποχική αύξηση των καμήλων γέννησης. Από τις 27 Αυγούστου 2014, υπήρξαν συνολικά 855 περιπτώσεις MERS-CoV, με 333 θανάτους και ποσοστό θνησιμότητας περίπου 40%, σύμφωνα με το Ευρωπαϊκό Κέντρο Πρόληψης και Ελέγχου Νόσων.
Το MERS-CoV είναι ένας β-κοροναϊός της ομάδας 2c που σχετίζεται σε μεγάλο βαθμό με δύο προηγουμένως αναγνωρισμένους κορωναϊούς, HKU4 και HKU5 [ 95 ]. Πιστεύεται ότι ο ιός προέρχεται από νυχτερίδες, αλλά πιθανότατα είχε έναν ενδιάμεσο ξενιστή, καθώς οι άνθρωποι σπάνια έρχονται σε επαφή με τη μυστική νυχτερίδα. Ορολογικές μελέτες έχουν εντοπίσει αντισώματα MERS-CoV σε καμήλες dromedary στη Μέση Ανατολή [ 96 ] και κυτταρικές σειρές από καμήλες έχουν βρεθεί ότι είναι επιτρεπτικές για την αντιγραφή MERS-CoV [ 97 ] παρέχοντας στοιχεία ότι οι καμήλες dromedary μπορεί να είναι ο φυσικός ξενιστής. Πιο πειστικές αποδείξεις προέρχονται από πρόσφατες μελέτες που εντοπίζουν σχεδόν ταυτόσημους MERS-CoVs τόσο σε καμήλες όσο και σε ανθρώπινες περιπτώσεις σε γειτονικές περιοχές της Σαουδικής Αραβίας [ 98 , 99]. Σε μία από αυτές τις μελέτες, η ανθρώπινη περίπτωση είχε άμεση επαφή με μολυσμένη καμήλα και ο ιός που απομονώθηκε από αυτόν ήταν πανομοιότυπος με τον ιό που απομονώθηκε από την καμήλα [ 99 ]. Επί του παρόντος παραμένει να καθοριστεί πόσες περιπτώσεις MERS-CoV μπορούν να αποδοθούν σε έναν ενδιάμεσο ξενιστή σε αντίθεση με τη μετάδοση από άνθρωπο σε άνθρωπο. Έχει επίσης υποτεθεί ότι η εξάπλωση από άνθρωπο σε καμήλα συνέβαλε στην έκρηξη.
Το MERS-CoV χρησιμοποιεί τη διπεπτιδυλική πεπτιδάση 4 (DPP4) ως υποδοχέα [ 100 ]. Ο ιός μπορεί να χρησιμοποιήσει μόνο τον υποδοχέα από ορισμένα είδη όπως νυχτερίδες, ανθρώπους, καμήλες, κουνέλια και άλογα για να διαπιστώσει μόλυνση. Δυστυχώς για τους ερευνητές, ο ιός δεν είναι σε θέση να μολύνει τα κύτταρα ποντικού λόγω διαφορών στη δομή της DPP4, καθιστώντας δύσκολη την αξιολόγηση πιθανών εμβολίων ή αντιιικών. Πρόσφατα, αναπτύχθηκε ένα μικρό ζωικό μοντέλο για MERS-CoV χρησιμοποιώντας έναν αδενοϊικό φορέα για να εισαχθεί το ανθρώπινο γονίδιο DPP4 σε πνεύμονες ποντικού [ 101 ]. Αυτό το μοναδικό σύστημα καθιστά δυνατή τη δοκιμή θεραπευτικών παρεμβάσεων και νέων εμβολίων για MERS-CoV σε οποιοδήποτε ζώο ευαίσθητο στις μεταβολές αδενοϊού.
Στις περισσότερες περιπτώσεις αυτοπεριορισμένης μόλυνσης, η διάγνωση των κοροναϊών είναι περιττή, καθώς η ασθένεια φυσικά θα διαρκέσει. Εντούτοις, μπορεί να είναι σημαντικό σε ορισμένες κλινικές και κτηνιατρικές εγκαταστάσεις ή σε επιδημιολογικές μελέτες να εντοπιστεί ένας αιτιολογικός παράγοντας. Η διάγνωση είναι επίσης σημαντική σε περιοχές όπου εμφανίζεται μια σοβαρή επιδημία του CoV, όπως επί του παρόντος στη Μέση Ανατολή, όπου η MERS-CoV εξακολουθεί να κυκλοφορεί. Ο προσδιορισμός των περιπτώσεων θα καθοδηγήσει την ανάπτυξη μέτρων δημόσιας υγείας για τον έλεγχο των εστιών. Είναι επίσης σημαντικό να εντοπιστούν περιπτώσεις σοβαρής κτηνιατρικής νόσου που προκαλείται από τον CoV, όπως οι PEDV και IBV, για τον έλεγχο αυτών των παθογόνων και την προστασία των τροφίμων. Η RT-PCR έχει γίνει η μέθοδος επιλογής για τη διάγνωση του ανθρώπινου CoV, καθώς έχουν αναπτυχθεί πολλαπλές δοκιμασίες RT-PCR σε πραγματικό χρόνο,102 ,103 ]. Οι ορολογικές δοκιμασίες είναι σημαντικές σε περιπτώσεις όπου το RNA είναι δύσκολο να απομονωθεί ή δεν υπάρχει πλέον, και για επιδημιολογικές μελέτες.
Μέχρι σήμερα, δεν υπάρχουν αντιιικά θεραπευτικά που να στοχεύουν ειδικά τους ανθρώπινους κοροναϊούς, έτσι οι θεραπείες είναι μόνο υποστηρικτικές. In vitro, οι ιντερφερόνες (IFNs) είναι μόνο εν μέρει αποτελεσματικές έναντι των κοροναϊών [ 104 ]. Οι IFNs σε συνδυασμό με ριμπαβιρίνη μπορεί να έχουν αυξημένη δραστικότητα in vitro σε σύγκριση με μόνο IFNs έναντι ορισμένων κοροναϊών. Ωστόσο, η αποτελεσματικότητα αυτού του συνδυασμού in vivo απαιτεί περαιτέρω αξιολόγηση [ 105 ]. Οι εστίες SARS και MERS έχουν διεγείρει την έρευνα σχετικά με αυτούς τους ιούς και αυτή η έρευνα έχει εντοπίσει μεγάλο αριθμό κατάλληλων αντιϊκών στόχων, όπως ιικές πρωτεάσες, πολυμεράσες και πρωτεΐνες εισόδου. Σημαντική δουλειά παραμένει, ωστόσο, η ανάπτυξη φαρμάκων που στοχεύουν σε αυτές τις διεργασίες και είναι σε θέση να αναστέλλουν την αναπαραγωγή του ιού.
Υπάρχουν μόνο περιορισμένες επιλογές για την πρόληψη λοιμώξεων από κορωναϊό. Τα εμβόλια έχουν εγκριθεί μόνο για τα IBV, TGEV και Canine CoV, αλλά αυτά τα εμβόλια δεν χρησιμοποιούνται πάντοτε επειδή είτε δεν είναι πολύ αποτελεσματικά είτε σε μερικές περιπτώσεις έχει αναφερθεί ότι εμπλέκονται στην επιλογή νέων παθογόνων CoV μέσω ανασυνδυασμού κυκλοφορούντων στελεχών. Τα εμβόλια για κτηνιατρικά παθογόνα, όπως το PEDV, μπορεί να είναι χρήσιμα σε περιπτώσεις όπου η εξάπλωση του ιού σε νέα θέση θα μπορούσε να οδηγήσει σε σοβαρές απώλειες κτηνιατρικών ζώων. Στην περίπτωση του SARS-CoV, έχουν αναπτυχθεί αρκετά πιθανά εμβόλια, αλλά δεν έχουν ακόμη εγκριθεί για χρήση. Αυτά τα εμβόλια περιλαμβάνουν ανασυνδυασμένους εξασθενημένους ιούς, ζωντανούς ιικούς φορείς ή μεμονωμένες ιικές πρωτεΐνες που εκφράζονται από πλασμίδια ϋΝΑ. Τα θεραπευτικά αντισώματα εξουδετέρωσης του SARS-CoV έχουν παραχθεί και θα μπορούσαν να ανακτηθούν και να χρησιμοποιηθούν και πάλι σε περίπτωση εμφάνισης άλλης εστίας SARS-CoV. Τέτοια αντισώματα θα ήταν πολύ χρήσιμα για την προστασία των εργαζομένων στον τομέα της υγείας. Γενικά, πιστεύεται ότι τα ζωντανά εξασθενημένα εμβόλια θα είναι τα πιο αποτελεσματικά στην στόχευση κοροναϊών. Αυτό απεδείχθη στην περίπτωση της TGEV, όπου στην Ευρώπη παρατηρήθηκε μια εξασθενημένη παραλλαγή, PRCV, στη δεκαετία του '80. Αυτή η παραλλαγή προκάλεσε μόνο ήπια ασθένεια και εντελώς προστατευμένους χοίρους από το TGEV. Έτσι, αυτός ο εξασθενημένος ιός προκάλεσε φυσικά την επανεμφάνιση σοβαρών TGEV στην Ευρώπη και τις ΗΠΑ τα τελευταία 30 χρόνια [ πιστεύεται ότι τα ζώντα εξασθενημένα εμβόλια θα είναι τα πιο αποτελεσματικά στην στόχευση των κοροναϊών. Αυτό απεδείχθη στην περίπτωση της TGEV, όπου στην Ευρώπη παρατηρήθηκε μια εξασθενημένη παραλλαγή, PRCV, στη δεκαετία του '80. Αυτή η παραλλαγή προκάλεσε μόνο ήπια ασθένεια και εντελώς προστατευμένους χοίρους από το TGEV. Έτσι, αυτός ο εξασθενημένος ιός προκάλεσε φυσικά την επανεμφάνιση σοβαρών TGEV στην Ευρώπη και τις ΗΠΑ τα τελευταία 30 χρόνια [ πιστεύεται ότι τα ζώντα εξασθενημένα εμβόλια θα είναι τα πιο αποτελεσματικά στην στόχευση των κοροναϊών. Αυτό απεδείχθη στην περίπτωση της TGEV, όπου στην Ευρώπη παρατηρήθηκε μια εξασθενημένη παραλλαγή, PRCV, στη δεκαετία του '80. Αυτή η παραλλαγή προκάλεσε μόνο ήπια ασθένεια και εντελώς προστατευμένους χοίρους από το TGEV. Έτσι, αυτός ο εξασθενημένος ιός προκάλεσε φυσικά την επανεμφάνιση σοβαρών TGEV στην Ευρώπη και τις ΗΠΑ τα τελευταία 30 χρόνια [106 ]. Παρά την επιτυχία αυτή, η ανάπτυξη εμβολίων για κοροναϊούς αντιμετωπίζει πολλές προκλήσεις [107 ]. Πρώτον, για λοιμώξεις του βλεννογόνου, η φυσική μόλυνση δεν εμποδίζει την επακόλουθη μόλυνση και έτσι τα εμβόλια πρέπει είτε να προκαλούν καλύτερη ανοσία από τον αρχικό ιό είτε πρέπει να ελαχιστοποιούν τουλάχιστον τη νόσο που εμφανίζεται κατά τη διάρκεια μιας δευτερογενούς λοίμωξης. Δεύτερον, η τάση των ιών να ανασυνδυασθούν μπορεί να αποτελέσει πρόβλημα καθιστώντας το εμβόλιο άχρηστο και αυξάνοντας ενδεχομένως την εξέλιξη και την ποικιλομορφία του ιού στην άγρια φύση [108 ]. Τέλος, έχει αποδειχθεί στο FIPV ότι ο εμβολιασμός με πρωτεΐνη S οδηγεί σε αυξημένη ασθένεια [109]. Παρόλα αυτά, αναπτύσσονται αρκετές στρατηγικές για την ανάπτυξη εμβολίου για να μειωθεί η πιθανότητα ανασυνδυασμού, για παράδειγμα κάνοντας μεγάλες διαγραφές στις πρωτεΐνες nsp1 [ 110 ] ή Ε [ 111 ], αναδιατάσσοντας το 3 'άκρο του γονιδιώματος [ 112 ] τις ακολουθίες TRS [ 113 ], ή χρησιμοποιώντας μεταλλαγμένους ιούς με μη φυσιολογικά υψηλά ποσοστά μετάλλαξης που εξασθενίζουν σημαντικά τον ιό [ 114 ].
Λόγω της έλλειψης αποτελεσματικών θεραπευτικών ή εμβολίων, τα καλύτερα μέτρα για τον έλεγχο των ανθρώπινων κοροναϊών παραμένουν ένα ισχυρό σύστημα επιτήρησης της δημόσιας υγείας, σε συνδυασμό με ταχείες διαγνωστικές εξετάσεις και απομόνωση όταν είναι απαραίτητο. Για τις διεθνείς εστίες, η συνεργασία κυβερνητικών φορέων, αρχών δημόσιας υγείας και παρόχων υγειονομικής περίθαλψης είναι κρίσιμη. Κατά τη διάρκεια κτηνιατρικών εκδηλώσεων που μεταδίδονται εύκολα, όπως το PEDV, ενδέχεται να χρειαστούν πιο δραστικά μέτρα, όπως η καταστροφή ολόκληρων κοπαδιών χοίρων, προκειμένου να αποτραπεί η μετάδοση αυτών των θανατηφόρων ιών.
Τα τελευταία 50 χρόνια, έχει εμφανιστεί η εμφάνιση πολλών διαφορετικών κοροναϊών που προκαλούν μεγάλη ποικιλία ανθρώπινων και κτηνιατρικών ασθενειών. Είναι πιθανό ότι αυτοί οι ιοί θα συνεχίσουν να εμφανίζονται και να εξελίσσονται και να προκαλούν τόσο ανθρώπινες όσο και κτηνιατρικές εστίες εξαιτίας της ικανότητάς τους να ανασυνδυάζουν, να μεταλλάσσουν και να μολύνουν πολλαπλούς τύπους και κυτταρικούς τύπους.
Η μελλοντική έρευνα για τους κοροναϊούς θα συνεχίσει να διερευνά πολλές πτυχές της αναπαραγωγής και παθογένεσης του ιού. Πρώτον, η κατανόηση της τάσης αυτών των ιών να πηδήξουν μεταξύ των ειδών, να διαπιστωθεί η μόλυνση σε έναν νέο ξενιστή και να εντοπιστούν σημαντικές δεξαμενές κοροναϊών θα βοηθήσει δραματικά στην ικανότητά μας να προβλέπουμε πότε και πού θα εμφανιστούν πιθανές επιδημίες. Δεδομένου ότι οι νυχτερίδες φαίνεται να είναι σημαντική δεξαμενή για τους ιούς αυτούς, θα είναι ενδιαφέρον να προσδιοριστεί πώς φαίνεται να αποφεύγουν κλινικά εμφανείς ασθένειες και να μολυνθούν επίμονα. Δεύτερον, πολλές από τις μη δομικές και βοηθητικές πρωτεΐνες που κωδικοποιούνται από αυτούς τους ιούς παραμένουν μη χαρακτηρισμένες με καμία γνωστή λειτουργία και θα είναι σημαντικό να αναγνωριστούν μηχανισμοί δράσης για αυτές τις πρωτεΐνες καθώς επίσης και ο ρόλος τους στον ιικό αναδιπλασιασμό και την παθογένεση. Αυτές οι μελέτες θα πρέπει να οδηγήσουν σε μεγάλη αύξηση του αριθμού των κατάλληλων θεραπευτικών στόχων για την καταπολέμηση λοιμώξεων. Επιπλέον, πολλά από τα μοναδικά ένζυμα που κωδικοποιούνται από κοροναϊούς, όπως η ADP-ριβόζη-1 "-φωσφατάση, υπάρχουν επίσης σε υψηλούς ευκαρυώτες, καθιστώντας τη μελέτη τους σχετική με την κατανόηση των γενικών πτυχών της μοριακής βιολογίας και της βιοχημείας. Τρίτον, η απόκτηση μιας ολοκληρωμένης εικόνας των περιπλοκών του RTC θα παρέχει ένα πλαίσιο για την κατανόηση της μοναδικής διαδικασίας αντιγραφής του RNA που χρησιμοποιείται από αυτούς τους ιούς. Τέλος, ο καθορισμός του μηχανισμού του τρόπου με τον οποίο οι κοροναϊοί προκαλούν ασθένεια και η κατανόηση της ανοσοπαθολογικής απόκρισης του ξενιστή θα βελτιώσει σημαντικά την ικανότητά μας να σχεδιάζουμε εμβόλια και να μειώνουμε την επιβάρυνση της νόσου. πολλά από τα μοναδικά ένζυμα που κωδικοποιούνται από κοροναϊούς, όπως η ADP-ριβόζη-1 "-φωσφατάση, είναι επίσης παρόντα σε υψηλούς ευκαρυώτες, καθιστώντας τη μελέτη τους σχετική με την κατανόηση των γενικών πτυχών της μοριακής βιολογίας και της βιοχημείας. Τρίτον, η απόκτηση μιας ολοκληρωμένης εικόνας των περιπλοκών του RTC θα παρέχει ένα πλαίσιο για την κατανόηση της μοναδικής διαδικασίας αντιγραφής του RNA που χρησιμοποιείται από αυτούς τους ιούς. Τέλος, ο καθορισμός του μηχανισμού του τρόπου με τον οποίο οι κοροναϊοί προκαλούν ασθένεια και η κατανόηση της ανοσοπαθολογικής απόκρισης του ξενιστή θα βελτιώσει σημαντικά την ικανότητά μας να σχεδιάζουμε εμβόλια και να μειώνουμε την επιβάρυνση της νόσου. πολλά από τα μοναδικά ένζυμα που κωδικοποιούνται από κοροναϊούς, όπως η ADP-ριβόζη-1 "-φωσφατάση, είναι επίσης παρόντα σε υψηλούς ευκαρυώτες, καθιστώντας τη μελέτη τους σχετική με την κατανόηση των γενικών πτυχών της μοριακής βιολογίας και της βιοχημείας. Τρίτον, η απόκτηση μιας ολοκληρωμένης εικόνας των περιπλοκών του RTC θα παρέχει ένα πλαίσιο για την κατανόηση της μοναδικής διαδικασίας αντιγραφής του RNA που χρησιμοποιείται από αυτούς τους ιούς. Τέλος, ο καθορισμός του μηχανισμού του τρόπου με τον οποίο οι κοροναϊοί προκαλούν ασθένεια και η κατανόηση της ανοσοπαθολογικής απόκρισης του ξενιστή θα βελτιώσει σημαντικά την ικανότητά μας να σχεδιάζουμε εμβόλια και να μειώνουμε την επιβάρυνση της νόσου. η απόκτηση μιας πλήρους εικόνας των περιπλοκών του RTC θα παρέχει ένα πλαίσιο για την κατανόηση της μοναδικής διαδικασίας αντιγραφής του RNA που χρησιμοποιείται από αυτούς τους ιούς. Τέλος, ο καθορισμός του μηχανισμού του τρόπου με τον οποίο οι κοροναϊοί προκαλούν ασθένεια και η κατανόηση της ανοσοπαθολογικής απόκρισης του ξενιστή θα βελτιώσει σημαντικά την ικανότητά μας να σχεδιάζουμε εμβόλια και να μειώνουμε την επιβάρυνση της νόσου. η απόκτηση μιας πλήρους εικόνας των περιπλοκών του RTC θα παρέχει ένα πλαίσιο για την κατανόηση της μοναδικής διαδικασίας αντιγραφής του RNA που χρησιμοποιείται από αυτούς τους ιούς. Τέλος, ο καθορισμός του μηχανισμού του τρόπου με τον οποίο οι κοροναϊοί προκαλούν ασθένεια και η κατανόηση της ανοσοπαθολογικής απόκρισης του ξενιστή θα βελτιώσει σημαντικά την ικανότητά μας να σχεδιάζουμε εμβόλια και να μειώνουμε την επιβάρυνση της νόσου.
1.
(2012) Ανταγωνισμός της οδού OAS-RNase L που προκαλείται από ιντερφερόνη από πρωτεΐνη ns2 από κορωναϊό ποντικού απαιτείται για τον αναδιπλασιασμό του ιού και την παθολογία του ήπατος. Microbe Host Cell 11: 607-616. doi: 10.1016 / j.chom.2012.04.011
2.
Barcena M, Oostergetel GT, Bartelink W et al (2009) Κρυοηλεκτρονική τομογραφία του ιού της ηπατίτιδας ποντικού: στοιχεία για τη δομή του coronavirion. Proc Natl Acad Sci USA 106: 582-587
3.
Neuman BW, Adair BD, Yoshioka C et al (2006) Η υπερμοριακή αρχιτεκτονική του κοροναϊού του σοβαρού οξέος αναπνευστικού συνδρόμου αποκαλυπτόμενη με κρυομικροσκοπία ηλεκτρονίων. J Virol 80: 7918 - 7928
4.
Beniac DR, Andonov Α, Grudeski E et al (2006) Αρχιτεκτονική της αιχμής της προφαρμάκου κορωνοαϊού SARS. Nat Struct Μοί ΒίοΙ 13: 751-752. doi: 10.1038 / nsmb1123
5.
Delmas B, Laude Η (1990) Συγκέντρωση της πρωτεΐνης αιχμής κοροναϊού σε τριμερή και ο ρόλος της στην έκφραση επιτόπου. J Virol 64: 5367 - 5375
6.
(2003) Η πρωτεΐνη spike coronavirus είναι πρωτεΐνη σύντηξης ιού κατηγορίας Ι: δομικό και λειτουργικό χαρακτηριστικό του συμπλόκου πυρήνα σύντηξης. J Virol 77: 8801 - 8811
7.
Collins AR, Knobler RL, Powell Η et αϊ (1982) Μονοκλωνικά αντισώματα στον ιό ηπατίτιδας-4 ποντικού (στέλεχος JHM) ορίζουν την ιική γλυκοπρωτεΐνη υπεύθυνη για πρόσδεση και σύντηξη κυττάρου-κυττάρου. Virology 119: 358-371
8.
Abraham S, Kienzle ΤΕ, Lapps W et αϊ (1990) Αφαίρεση της αλληλουχίας της πρωτεΐνης αιχμής κορώνα βοοειδούς και ταυτοποίηση της εσωτερικής πρωτεολυτικής θέσης διάσπασης. Virology 176: 296-301
9.
Luytjes W, Sturman LS, Bredenbeek PJ et αϊ. (1987) Πρωτεύουσα δομή της γλυκοπρωτεΐνης Ε2 του κορανοϊού MHV-A59 και ταυτοποίηση της θέσης διάσπασης θρυψίνης. Virology 161: 479-487
10.
de Groot RJ, Luytjes W, Horzinek MC et αϊ (1987) Στοιχεία για δομή περιέλιξης-πηνίου στις πρωτεΐνες αιχμής των κοροναϊών. J Mol Biol 196: 963-966
11.
Armstrong J, Niemann Η, Smeekens δ et αϊ (1984) Αλληλουχία και τοπολογία μίας πρότυπης πρωτεΐνης ενδοκυτταρικής μεμβράνης, ΕΙ γλυκοπρωτεΐνης, από κοροναϊό. Nature 308: 751-752
12.
Nal B, Chan C, Kien F et al (2005) Διαφορική ωρίμανση και υποκυτταρικός εντοπισμός σοβαρών επιφανειακών πρωτεϊνών επιφανείας κοροναϊού σοβαρής αναπνευστικού συνδρόμου S, Μ και Ε. Gen. Virol 86: 1423-1434. doi: 10.1099 / vir.0.80671-0
13.
Neuman BW, Kiss G, Kunding AH et al (2011) Μια δομική ανάλυση της πρωτεΐνης Μ στην συναρμολόγηση και τη μορφολογία του κοροναϊού. J Struct Biol 174: 11-22. doi: 10.1016 / j.jsb.2010.11.021
14.
Ο Godet Μ, L'Haridon R, Vautherot JF et αϊ (1992) ο TGEV ιός corona ORF4 κωδικοποιεί μία μεμβρανική πρωτεΐνη που ενσωματώνεται σε βιριόνια. Virology 188: 666-675
15.
DeDiego ML, Alvarez E, Almazan F et al (2007) Ένα σοβαρό οξείδιο του αναπνευστικού συνδρόμου κοροναϊό που στερείται του γονιδίου Ε εξασθενίζει in vitro και in vivo. J Virol 81: 1701-1713
16.
Nieto-Torres JL, Dediego ML, Verdia-Baguena C et al (2014) Η δραστικότητα του καναλιού ιόντων της πρωτεΐνης του κοροναϊού σοβαρής οξείας αναπνευστικής συνείδησης προάγει την καταλληλότητα του ιού και την παθογένεση. PLoS Pathog 10: e1004077. doi: 10.1371 / journal.ppat.1004077
17.
Chang CK, Sue SC, Yu TH et al (2006) Modular οργάνωση της νουκλεοκαψιδικής πρωτεΐνης του coronavirus. J Biomed Sci 13: 59-72. doi: 10.1007 / s11373-005-9035-9
18.
Hurst KR, Koetzner CA, Masters PS (2009) Ταυτοποίηση in νίνο-αλληλεπιδρώντων περιοχών της νουκλεοκαψιδικής πρωτεΐνης μυϊκού κορώνα ιού. J Virol 83: 7221-7234. doi: 10.1128 / JVI.00440-09
19.
Stohlman SA, Lai ΜΜ (1979) Φωσφοπρωτεΐνες ιού ηπατίτιδας ποντικού. J Virol 32: 672-675
20.
Stohlman SA, Baric RS, Nelson GN et αϊ (1988) Ειδική αλληλεπίδραση μεταξύ RNA οδηγού κοροναϊού και πρωτεΐνης νουκλεοκαψιδίου. J Virol 62: 4288 - 4295
21.
Molenkamp R, Spaan WJ (1997) Ταυτοποίηση μιας ειδικής αλληλεπίδρασης μεταξύ της νουκλεοκαψιδικής πρωτεΐνης A59 του ιού της ηπατίτιδας ποντικού κοροναϊού και του σήματος συσκευασίας. Virology 239: 78-86
Hurst KR, Koetzner CA, Masters PS (2013) Χαρακτηρισμός μιας κρίσιμης αλληλεπίδρασης μεταξύ της πρωτεΐνης νουκλεοκαψιδίου κοροναϊού και της μη δομικής πρωτεΐνης 3 του συμπλόκου της ιικής αντιγραφάσης-μεταγραφάσης. J Virol 87: 9159-9172. doi: 10.1128 / JVI.01275-13
24.
Sturman LS, Holmes KV, Behnke J (1980) Απομόνωση γλυκοπρωτεϊνών περιβλήματος κοροναϊού και αλληλεπίδραση με το νουκλεοκαψίδιο. J Virol 33: 449 - 462
25.
Klausegger Α, Strobl Β, Regl G et al (1999) Ταυτοποίηση μιας αιμογλουτινίνης-εστεράσης κορωνοϊού με ειδικότητα υποστρώματος διαφορετικό από εκείνο του ιού της γρίπης C και του κορωναϊού των βοοειδών. J Virol 73: 3737-3743
26.
Cornelissen LA, Wierda CM, van der Meer FJ et αϊ (1997) Αιμαγλουτινίνη-εστεράση, νέα δομική πρωτεΐνη τουτοϊού. J Virol 71: 5277 με 5286
27.
Kazi L, Lissenberg A, Watson R et al (2005) Η έκφραση της πρωτεΐνης εστεράσης αιμασυσσωματίνης από τον ανασυνδυασμένο ιό της ηπατίτιδας ποντικού ενισχύει τη νευροβιολογική δράση. J Virol 79: 15064 με 15073
28.
Lissenberg Α, Vrolijk MM, van Vliet AL et al (2005) Luxury με κόστος; Οι ανασυνδυασμένοι ιοί ηπατίτιδας ποντικού που εκφράζουν την βοηθητική πρωτεΐνη εστεροειδούς αιμοσυγκολλητίνης μειώνουν τη φυσική κατάσταση in vitro. J Virol 79: 15054 με 15.063
29.
Kubo Η, Yamada ΥΚ, Taguchi F (1994) Τοποθέτηση εξουδετερωτικών επιτόπων και της θέσης δέσμευσης υποδοχέα εντός των αμινοτερματικών 330 αμινοξέων της πρωτεΐνης αιχμής κορώνα ιού μυοειδούς. J Virol 68: 5403 - 5.410
30.
Cheng PK, Wong DA, Tong LK και συνεργάτες (2004) Πρότυπα ιογενής απόσπασης του κοροναϊού σε ασθενείς με πιθανό σοβαρό οξύ αναπνευστικό σύνδρομο. Lancet 363: 1699-1700
31.
Belouzard S, Chu VC, Whittaker GR (2009) Ενεργοποίηση της πρωτεΐνης ακανθοκυττάρων SARS μέσω διαδοχικής πρωτεολυτικής διάσπασης σε δύο ξεχωριστές θέσεις. Proc Natl Acad Sci USA 106: 5871-5876. doi: 10.1073 / pnas.0809524106
32.
Baranov PV, Henderson CM, Anderson CB et al (2005) Προγραμματισμένη ριζοσωματική μετατόπιση πλαισίου στην αποκωδικοποίηση του γονιδιώματος SARS-CoV. Virology 332: 498-510. doi: 10.1016 / j.virol.2004.11.038
(2010) Τα παθογόνα ειδικά για τον ιό Τ κύτταρα προκαλούν ασθένεια κατά τη διάρκεια της θεραπείας με τον αναστολέα καλσινευρίνης FK506: επιπλοκές για τη μεταμόσχευση. J Exp Med 207: 2355-2367
35.
Ziebuhr J, Snijder EJ, Gorbalenya ΑΕ (2000) Κινάσες που κωδικοποιούνται από ιούς και πρωτεολυτική επεξεργασία στα Nidovirales. J Gen Virol 81: 853-879
36.
Mielech AM, Chen Y, Mesecar AD et al (2014) Nidovirus πρωτεΐνες τύπου παπαϊνης: πολυλειτουργικά ένζυμα με δραστικότητα πρωτεάσης, αποβουκικιτίνης και αποσυλλεργίας. Virus Res. doi: 10.1016 / j.virusres.2014.01.025
37.
Snijder EJ, Bredenbeek PJ, Dobbe JC et al (2003) Μοναδικά και συντηρημένα χαρακτηριστικά του γονιδιώματος και του πρωτεόνος του SARS-coronavirus, ένα πρώιμο αποχωρισμό από την ομάδα της σειράς 2 του coronavirus. J Mol Biol 331: 991-1004
38.
Sethna PB, Hofmann ΜΑ, Brian DA (1991) Τα αντίγραφα με ελάχιστη έλικα των αντιγραφόντων mRNA των κορωνοϊών περιέχουν αντικαταθλιπτικά. J Virol 65: 320-325
39.
(2007) Ένας βρόχος βλαστοκυττάρων RNA εντός της κωδικεύουσας περιοχής nsp1 βοοειδούς κοροναϊού είναι ένα cis-ενεργό στοιχείο στην ελαττωματική αναπαραγωγή RNA παρεμβολής. J Virol 81: 7716-7724. doi: 10.1128 / JVI.00549-07
40.
Ο βέλτιστος cis-αναδιπλασιασμός βλαστικού βρόχου IV στην 5 'αμετάφραστη περιοχή του γονιδιώματος του κορωναϊού ποντικού εκτείνεται 16 νουκλεοτίδια σε ανοιχτό πλαίσιο ανάγνωσης 1. J Virol 85: 5593-5605. doi: 10.1128 / JVI.00263-11
41.
Ο Liu P, Li L, Keane SC et al (2009) Ο βλαστικός βρόχος 2 του ιού της ηπατίτιδας του ποντικού υιοθετεί μία δομή tetraloop uYNMG (U) α που είναι εξαιρετικά λειτουργική ανεκτική στις υποκαταστάσεις βάσης. J Virol 83: 12084-12093. doi: 10.1128 / JVI.00915-09
42.
Οι Raman S, Bouma Ρ, Williams GD et αϊ (2003) Stem-loop III στην μη μεταφρασμένη περιοχή 5 'είναι ένα cis-ενεργό στοιχείο στην ελαττωματική αντιγραφή RNA αντιγραφής του βόειου κορωναϊού. J Virol 77: 6720 - 6730
43.
Liu Q, Johnson RF, Leibowitz JL (2001) Δευτερεύοντα δομικά στοιχεία εντός της 3 'μη μεταφρασμένης περιοχής του γονιδιωματικού RNA του ιού της ηπατίτιδας JHM ποντικού. J Virol 75: 12105-12113. doi: 10.1128 / JVI.75.24.12105-12113.2001
44.
(2007) Μια υπερμεταβλητή περιοχή εντός του 3 'cis-ενεργού στοιχείου του γονιδιώματος του κορωναϊού ποντικού δεν είναι απαραίτητη για τη σύνθεση του RNA αλλά επηρεάζει την παθογένεση. J Virol 81: 1274-1287. doi: 10.1128 / JVI.00803-06
45.
Williams GD, Chang RY, Brian DA (1999) Μια φυλλογενετικώς συντηρημένη 3 'αμετάφραστη περιοχή pseudoknot τύπου 3' μη-μεταφρασμένης περιοχής λειτουργεί σε αναδιπλασιασμό RNA κορωνοϊού. J Virol 73: 8349-8355
46.
Hsue Β, Masters PS (1997) Μια διόγκωση δομή βρόχου-βρόχου στην 3 'αμετάφραστη περιοχή του γονιδιώματος του ιού ηπατίτιδας ποντικού κοροναϊού είναι απαραίτητη για την αντιγραφή. J Virol 71: 7567-7578
47.
Hsue Β, Hartshorne Τ, Masters PS (2000) Χαρακτηρισμός μιας δευτερεύουσας δομής βασικού RNA στην 3 'αμετάφραστη περιοχή του γονιδιώματος του κορωναϊού ποντικού. J Virol 74: 6911-6921
48.
Sawicki SG, Sawicki DL, Siddell SG (2007) Μια σύγχρονη άποψη της μεταγραφής του κοροναϊού. J Virol 81: 20-29
49.
Bentley K, Keep SM, Armesto M et al (2013) Ταυτοποίηση ενός μη κανανικά μεταγεγραμμένου υπογονιδιωματικού mRNA ιού μολυσματικής βρογχίτιδας και άλλων γαμακορονικών ιικών. J Virol 87: 2128-2136. doi: 10.1128 / JVI.02967-12
50.
Keck JG, Makino S, Soe LH et αϊ (1987) RNA ανασυνδυασμός κοροναϊού. Adv Εχρ. Med Biol 218: 99-107
51.
Lai ΜΜ, Baric RS, Makino δ et αϊ (1985) Ανασυνδυασμός μεταξύ γονιδιωμάτων RNA που δεν έχουν διαχωριστεί από κορωναϊούς ποντικού. J Virol 56: 449-456
52.
(1994) Χαρακτηρισμός του εκκολαπτόμενου τμήματος του ιού της ηπατίτιδας ποντικού: στοιχεία που αποδεικνύουν ότι η μεταφορά από το RER προς το σύμπλεγμα Golgi απαιτεί μόνο ένα στάδιο μεταφοράς των κυψελίδων. J Cell Biol 124: 55-70
53.
Tooze J, Tooze S, Warren Ο (1984) Replication of coronavirus MHV-A59 σε κύτταρα σάκκου: προσδιορισμός της πρώτης θέσης εκβλάστησης των απογόνων βιριόνων. Eur J Cell Βίο 33: 281-293
54.
de Haan CA, Rottier PJ (2005) Μοριακές αλληλεπιδράσεις στη συναρμολόγηση των κοροναϊών. Adv Virus Res 64: 165-230
55.
Bos EC, Luytjes W, van der Meulen HV et αϊ (1996) Η παραγωγή ανασυνδυασμένων μολυσματικών DI-σωματιδίων κορωναϊού μυοειδούς απουσία βοηθητικού ιού. Virology 218: 52-60
56.
(2008) Οι Μ, Ε και Ν δομικές πρωτεΐνες του κοροναϊού σοβαρής οξείας αναπνευστικού συνδρόμου απαιτούνται για αποτελεσματική συναρμολόγηση, διακίνηση και απελευθέρωση σωματιδίων που μοιάζουν με ιούς. J Virol 82: 11318-11330. doi: 10.1128 / JVI.01052-08
57.
Raamsman MJ, Locker JK, de Hooge Α et αϊ (2000) Χαρακτηρισμός της πρωτεΐνης μικρής μεμβράνης ιού ηπατίτιδας ποντικού κορωναϊού ποντικού A59 Ε. J Virol 74: 2333-2342
58.
Corse E, Machamer CE (2000) Η πρωτεΐνη Ε του ιού της λοιμώδους βρογχίτιδας στοχεύει στο σύμπλεγμα Golgi και κατευθύνει την απελευθέρωση σωματιδίων που μοιάζουν με ιούς. J Virol 74: 4319-4326
59.
Fischer F, Stegen CF, Masters PS et al (1998) Η ανάλυση κατασκευασμένων μεταλλαγμάτων γονιδίου Ε του ιού ηπατίτιδας ποντικού επιβεβαιώνει έναν κεντρικό ρόλο για την πρωτεΐνη Ε σε συναρμολόγηση κορωναϊού. J Virol 72: 7885 έως 7894
60.
Οι παλμιτοποιήσεις πρωτεϊνών φακέλου είναι ζωτικής σημασίας για τη συναρμολόγηση του κορωναϊού ποντικού. J Virol 82: 2989-2999. doi: 10.1128 / JVI.01906-07
61.
Ye Y, Hogue BG (2007) Ο ρόλος της διαμεμβρανικής περιοχής της πρωτεΐνης ιωοπροϊνης Ε κοροναϊού σε συναρμολόγηση ιού. J Virol 81: 3597-3607. doi: 10.1128 / JVI.01472-06
62.
Hurst KR, Kuo L, Koetzner CA et al (2005) Ένας κύριος καθοριστής για την αλληλεπίδραση πρωτεϊνών μεμβράνης εντοπίζεται στην καρβοξυ-τερματική περιοχή της πρωτεΐνης νουκλεοκαψιδίου του κορωναϊού ποντικού. J Virol 79: 13285-13297
63.
Perlman S, Netland J (2009) Coronaviruses post-SARS: ενημέρωση σχετικά με την αναπαραγωγή και την παθογένεση. Nat Rev Microbiol 7: 439-450
64.
Mihindukulasuriya KA, Wu G, St LJ et al (2008) Ταυτοποίηση ενός νέου κορωναϊού από μια φάλαινα beluga με τη χρήση μίας μικροσυστοιχίας πανοπλίας. J Virol 82: 5084 - 5088
65.
Ο B, Zhang Y, Xu L et al (2014) Ταυτοποίηση ποικίλων αλφακονοϊών και γονιδιωματικού χαρακτηρισμού ενός νέου βαριάς μορφής οξειδωμένου από το αναπνευστικό σύνδρομο κορωναϊού από νυχτερίδες στην Κίνα. J Virol 88: 7070-7082. doi: 10.1128 / JVI.00631-14
66.
Nga PT, Παρκέ Mdel C, Lauber C et al (2011) Ανακάλυψη του πρώτου νιδοϊού εντόμων, ελλειπόμενη εξελικτική σύνδεση στην εμφάνιση των μεγαλύτερων γονιδιωμάτων του ιού RNA. PLoS Pathog 7: e1002215. doi: 10.1371 / journal.ppat.1002215
67.
Lauber C, Ziebuhr J, Junglen S et al (2012) Mesoniviridae: μια προτεινόμενη νέα οικογένεια με τη σειρά Nidovirales που σχηματίζεται από ένα μόνο είδος ιού που μεταδίδεται από κουνουπιές. Arch Virol 157: 1623-1628. doi: 10.1007 / s00705-012-1295-x
68.
Levy GA, Liu Μ, Ding J et al (2000) Μοριακή και λειτουργική ανάλυση του ανθρώπινου γονιδίου προθρομβινάσης (HFGL2) και του ρόλου του στην ιική ηπατίτιδα. Am J Pathol 156: 1217-1225
69.
Lampert PW, Sims JK, Kniazeff AJ (1973) Μηχανισμός απομυελίνωσης στην εγκεφαλομυελίτιδα του ιού JHM. Acta Neuropathol 24: 76-85
70.
Weiner LP (1973) Παθογένεια απομυελίνωσης που προκαλείται από ιό ηπατίτιδας ποντικού (ιός JHM). Arch Neurol 28: 298-303
71.
Wu GF, Dandekar ΑΑ, Pewe L et αϊ (2000) Τα CD4 και CD8 Τ κύτταρα έχουν περιττούς αλλά όχι ταυτόσημους ρόλους σε απομυελίνωση που προκαλείται από ιό. J Immunol 165: 2278-2286
72.
Wang F, Stohlman SA, Fleming JO (1990) Διεγείρεται ανοσολογικώς η απομυελίνωση που επάγεται από στελέχη JHM ιού ηπατίτιδας μυοειδούς (MHV-4). J Neuroimmunol 30: 31-41
73.
Wu GF, Perlman S (1999) Η διείσδυση μακροφάγων, αλλά όχι η απόπτωση, συσχετίζεται με ανοσομεσολαβούμενη απομυελίνωση μετά από μόλυνση από μυϊκό ιστό με νευροτροπικό κοροναϊό. J Virol 73: 8771 - 8780
74.
McIntosh Κ, Becker WB, Chanock RM (1967) Ανάπτυξη στον εγκέφαλο ποντικού-ποντικού των ιών «τύπου ΙΒν» από ασθενείς με ασθένεια της ανώτερης αναπνευστικής οδού. Proc Natl Acad Sci USA 58: 2268-2273
75.
Bradburne AF, Bynoe ML, Tyrell DAJ (1967) Επιδράσεις ενός "νέου" ανθρώπινου αναπνευστικού ιού σε εθελοντές. Br Med 3: 767-769
76.
Hamre D, Procknow JJ (1966) Ένας νέος ιός που απομονώνεται από την ανθρώπινη αναπνευστική οδό. Proc Soc Exp ΒίοΙ Med 121: 190-193
77.
Woo PC, Lau SK, Chu CM et al (2005) Χαρακτηρισμός και πλήρης γονιδιωματική αλληλουχία ενός νέου coronavirus, coronavirus HKU1, από ασθενείς με πνευμονία. J Virol 79: 884-895
78.
van der Hoek L, Pyrc K, Jebbink MF et al (2004) Ταυτοποίηση ενός νέου ανθρώπινου κοροναϊού. Nat Med 10: 368-373
79.
van der Hoek L, Sure K, Ihorst G et al (2005) Η Croup συνδέεται με τον νέο κορωναϊό NL63. PLoS Med 2: E240
80.
Chibo D, Birch C (2006) Η ανάλυση των γονιδίων ακμής και νουκλεοπρωτεϊνών ανθρώπινου κοροναϊού 229E αποδεικνύει γενετική μετατόπιση μεταξύ χρονολογικά διακριτών στελεχών. J Gen Virol 87: 1203-1208
81.
Vijgen L, Keyaerts Ε, Lemey Ρ et al (2005) Κυκλοφορία γενετικά διακριτών σύγχρονων στελεχών OC43 ανθρώπινου κορωναϊού. Virology 337: 85-92
82.
Guan Y, Zheng BJ, He YQ et al (2003) Απομόνωση και χαρακτηρισμός ιών που σχετίζονται με τον κορανοϊό του SARS από ζώα στη νότια Κίνα. Science 302: 276-278
83.
Lau SK, Woo PC, Li KS et al (2005) Σοβαρές οξείες αναπνευστικές λοιμώξεις του ιού τύπου κοροναϊού σε κινέζικα νυχτερίδες. Proc Natl Acad Sci USA 102: 14040 έως 14.045
84.
Li W, Shi Z, Yu M et al (2005) Οι νυχτερίδες είναι φυσικές δεξαμενές κορωνοϊών τύπου SARS. Science 310: 676-679
85.
Ge XY, Li JL, Yang XL et al (2013) Απομόνωση και χαρακτηρισμός ενός coronavirus τύπου SARS που χρησιμοποιεί τον υποδοχέα ACE2. Nature 503: 535-538. doi: 10.1038 / nature12711
86.
Peiris JS, Yuen KY, Osterhaus AD et al (2003) Το σοβαρό οξύ αναπνευστικό σύνδρομο. N Engl J Med 349: 2431-2441
87.
Peiris JS, Chu CM, Cheng VC et al (2003) Κλινική εξέλιξη και ιϊκό φορτίο σε μια κοινοτική εστία πνευμονίας του SARS που σχετίζεται με κοροναϊό: μια προοπτική μελέτη. Lancet 361: 1767-1772
88.
Spiegel M, Schneider K, Weber F et al (2006) Αλληλεπίδραση του σοβαρού οξέος στεροειδούς αναπνευστικού συνδρόμου με τα δενδριτικά κύτταρα. J Gen Virol 87: 1953-1960
89.
Νόμος HK, Cheung CY, Ng ΗΥ et αϊ (2005) Ρυθμιστική ρύθμιση χημειοκίνης σε επιμολυσμένα από ανθρώπινα μονοκύτταρα δενδριτικά κύτταρα που έχουν μολυνθεί από SARS. Αίμα 106: 2366-2376
Roberts A, Paddock C, Vogel L et al (2005) Τα ηλικιωμένα BALB / c ποντίκια ως μοντέλο για αυξημένη σοβαρότητα σοβαρού οξέος αναπνευστικού συνδρόμου σε ηλικιωμένους ανθρώπους. J Virol 79: 5.833 έως 5838
92.
Zhao J, Zhao J, Perlman S (2010) Οι αποκρίσεις Τ κυττάρων απαιτούνται για την προστασία από την κλινική ασθένεια και για την κάθαρση ιού σε μολυσμένα με οξεία αναπνευστικό σύνδρομο μολυσμένα με κοροναϊό ποντίκια. J Virol 84: 9318-9325
93.
Οι σχετιζόμενες με την ηλικία αυξήσεις στην έκφραση PGD (2) μειώνουν την αναπνευστική μετανάστευση DC, με αποτέλεσμα την ελαττωμένη απόκριση των Τ κυττάρων κατά τη μόλυνση του αναπνευστικού ιού σε ποντίκια. J Clin Invest 121: 4921-4930. doi: 10.1172 / JCI59777
94.
Zaki AM, van Boheemen S, Bestebroer TM et al (2012) Απομόνωση ενός νέου κορωναϊού από έναν άνθρωπο με πνευμονία στη Σαουδική Αραβία. N Engl J Med 367: 1814-1820. doi: 10.1056 / NEJMoa1211721
95.
van Boheemen S, de Graaf M, Lauber C et al (2012) Γονιδιωματικός χαρακτηρισμός ενός πρόσφατα ανακαλυφθέντος κοροναϊού που σχετίζεται με σύνδρομο οξείας αναπνευστικής δυσφορίας στους ανθρώπους. MBio 3. doi: 10.1128 / mBio.00473-12
96.
Meyer Β, Muller ΜΑ, Corman VM κ.ά. (2014) Αντισώματα έναντι του MORES κοροναϊού σε καμήλες Dromedary, Ηνωμένα Αραβικά Εμιράτα, 2003 και 2013. Emerg Infect Dis 20: 552-559. doi: 10.3201 / eid2004.131746
97.
Eckerle I, Corman VM, Muller ΜΑ και άλλοι (2014) Αναπαραγωγική ικανότητα του MERS κοροναϊού σε κυτταρικές σειρές ζώων. Emerg Infect Dis 20: 276-279. doi: 10.3201 / eid2002.131182
98.
Memish ZA, Cotten M, Meyer Β et al (2014) Ανθρώπινη μόλυνση με MERS coronavirus μετά από έκθεση σε μολυσμένες καμήλες, Σαουδική Αραβία, 2013. Emerg Infect Dis 20: 1012-1015. doi: 10.3201 / eid2006.140402
99.
Azhar EI, El-Kafrawy SA, Farraj SA et al (2014) Στοιχεία για μετάδοση από καμήλα σε άνθρωπο του MERS coronavirus. N Engl J Med. doi: 10.1056 / NEJMoa1401505
100.
Raj VS, Mou Η, Smits SL et al (2013) Η διπεπτιδυλική πεπτιδάση 4 είναι ένας λειτουργικός υποδοχέας για το αναδυόμενο ανθρώπινο κορωναϊό-ΗΜC. Nature 495: 251-254. doi: 10.1038 / nature12005
101.
Zhao J, Li K, Wohlford-Lenane C et al (2014) Ταχεία παραγωγή ενός μοντέλου ποντικού για αναπνευστικό σύνδρομο στη Μέση Ανατολή. Proc Natl Acad Sci USA 111: 4970-4975. doi: 10.1073 / pnas.1323279111
102.
Emery SL, Erdman DD, Bowen MD et al (2004) Ανάλυση αλυσιδωτής αντίδρασης ανάστροφης μεταγραφής-πολυμεράσης σε πραγματικό χρόνο για κορωναϊό που σχετίζεται με SARS. Emerg Infect Dis 10: 311-316. doi: 10.3201 / eid1002.030759
103.
(2010) Η επιδημιολογία και οι κλινικές παρουσιάσεις των τεσσάρων ανθρώπινων κοροναϊών 229Ε, HKU1, NL63 και OC43 ανιχνεύθηκαν σε 3 χρόνια χρησιμοποιώντας μια νέα πολυδύναμη μέθοδο PCR πραγματικού χρόνου. J Clin Microbiol 48: 2940-2947. doi: 10.1128 / JCM.00636-10
104.
Cinatl J, Morgenstern Β, Bauer G et al (2003) Θεραπεία του SARS με ανθρώπινες ιντερφερόνες. Lancet 362: 293-294
105.
Stockman LJ, Bellamy R, Garner Ρ (2006) SARS: συστηματική ανασκόπηση των επιδράσεων της θεραπείας. PLoS Med 3: e343
106.
Laude Η, Van Reeth Κ, Pensaert Μ (1993) Κορνοϊός του αναπνευστικού του χοίρου: μοριακά χαρακτηριστικά και αλληλεπιδράσεις ιού-ξενιστή. Vet Res 24: 125-150
107.
Saif LJ (2004) Εμβόλια ζωϊκού κορωναϊού: μαθήματα για το SARS. Dev Biol (Βασιλεία) 119: 129-140
108.
Wang L, Junker D, Collisson EW (1993) Απόδειξη φυσικού ανασυνδυασμού στο γονίδιο S1 του ιού της μολυσματικής βρογχίτιδας. Virology 192: 710-716
109.
Vennema Η, de Groot RJ, Harbor DA et αϊ (1990) Πρώιμος θάνατος μετά από πρόκληση ιού μολυσματώδους περιτονίτιδας από αιλουροειδή λόγω ανοσοποίησης με ανασυνδυασμένο ιό δαμαλίτιδας. J Virol 64: 1407-1409
110.
Zust R, Cervantes-Barragan L, Kuri Τ et al (2007) Η μη διαρθρωτική πρωτεΐνη 1 του Coronavirus είναι ένας σημαντικός παράγοντας παθογονικότητας: επιπτώσεις για τον ορθολογικό σχεδιασμό των εμβολίων με κοροναϊό. PLoS Παθολογία 3: Ε109
111.
Ο Netonia J, DeDiego ML, Zhao J et al (2010) Η ανοσοποίηση με εξασθενημένο οξύ οξύ αναπνευστικό σύνδρομο κοροναϊό που διαγράφεται σε πρωτεΐνη Ε προστατεύει από θανατηφόρο αναπνευστικό νόσημα. Virology 399: 120-128. doi: 10.1016 / j.virol.2010.01.004
112.
de Haan CA, Volders Η, Koetzner CA κ.ά. (2002) Οι κοροναϊοί διατηρούν βιωσιμότητα παρά τις δραματικές αναδιατάξεις της αυστηρά διατηρημένης γονιδιακής οργάνωσης. J Virol 76: 12491 - 12.502
113.
Yount B, Roberts RS, Lindesmith L et al (2006) Επανασύνδεση του κυκλικού μεταγραφικού κυκλώματος οξείας αναπνευστικού συνδρόμου (SARS-CoV): σχεδίαση γονιδίου ανθεκτικού σε ανασυνδυασμό. Proc Natl Acad Sci USA 103: 12546 έως 12551
114.
Graham RL, Becker MM, Eckerle LD et al (2012) Ένα ζωντανό εμβόλιο κοροναϊού με μειωμένη εμπιστοσύνη προστατεύει σε ένα ηλικιωμένο, ανοσοκατασταλμένο μοντέλο ποντικού με θανατηφόρο νόσημα. Nat Med. 18: 1820-1826. doi: 10.1038 / nm.2972
115.
Yeager CL, Ashmun RA, Williams RK et αϊ (1992) Η ανθρώπινη αμινοπεπτιδάση Ν είναι ένας υποδοχέας για ανθρώπινο κοροναϊό 229Ε. Nature 357: 420-422. doi: 10.1038 / 357420a0
116.
Ο Hofmann H, Pyrc K, van der Hoek L et al (2005) Ο ανθρώπινος κορωναϊός NL63 χρησιμοποιεί τον υποδοχέα κοροναϊού για οξύ οξύ αναπνευστικό σύνδρομο για κυτταρική είσοδο. Proc Natl Acad Sci USA 102: 7988-7993
117.
Οι Delmas Β, Gelfi J, L'Haridon R et αϊ (1992) Η αμινοπεπτιδάση Ν είναι ένας κύριος υποδοχέας για τον εντερο-παθογόνο κοροναϊό TGEV. Nature 357: 417-420. doi: 10.1038 / 357417a0
118.
Li BX, Ge JW, Li YJ (2007) Η αμινοπεπτιδάση Ν του χοίρου Ν είναι ένας λειτουργικός υποδοχέας για τον κοροναϊό του PEDV. Virology 365: 166-172. doi: 10.1016 / j.virol.2007.03.031
119.
Tresnan DB, Levis R, Holmes KV (1996) Feline αμινοπεπτιδάση Ν χρησιμεύει ως υποδοχέας για αιλουροειδή, κυνοειδή, χοίρου, και ανθρώπινων coronaviruses σε οροομάδας I. J Virol 70: 8669-8674
120.
Benbacer L, Kut Ε., Besnardeau L et αϊ (1997) Οι χιμαιρικές αλληλουχίες αμινοπεπτιδάσης-ινδέρματος μεταξύ των ειδών αποκαλύπτουν αναγνώριση ειδικού υποδοχέα ειδών από κορωνοϊό σκύλου, ιό μολυσματικής περιτονίτιδας από αιλουροειδή και ιό μεταδοτικής γαστρεντερίτιδας. J Virol 71: 734-737
121.
Nedellec Ρ, Dveksler GS, Daniels Ε et αϊ (1994) Bgp2, ένα νέο μέλος της οικογένειας γονιδίων που σχετίζονται με καρκινοεμβρυονικό αντιγόνο, κωδικοποιεί έναν εναλλακτικό υποδοχέα για ιούς ηπατίτιδας ποντικού. J Virol 68: 4525 με 4537
122.
Williams RK, Jiang GS, Holmes KV (1991) Ο υποδοχέας για ιό ηπατίτιδας ποντικού είναι μέλος της οικογένειας των καρκινοεμβρυονικών αντιγόνων των γλυκοπρωτεϊνών. Proc Natl Acad Sci USA 88: 5533-5536
123.
Schultze Β, Herrler Ο (1992) Ο κοροναϊός των βοοειδών χρησιμοποιεί Ν-ακετυλο-9-Ο-ακετυλο-νευραμινικό οξύ ως καθοριστικό υποδοχέα για την έναρξη της μόλυνσης των καλλιεργημένων κυττάρων. J Gen Virol 73 (Pt 4): 901-906
124.
Li W, Moore MJ, Vasilieva Ν et al (2003) Το ένζυμο μετατροπής αγγειοτασίνης 2 είναι ένας λειτουργικός υποδοχέας για τον κορανοϊό του SARS. Nature 426: 450-454
125.
Η πρωτεΐνη nsp1 μεταδιδόμενου ιού της γαστρεντερίτιδας του Alphacoronavirus καταστέλλει την πρωτεϊνική μετάφραση σε κύτταρα θηλαστικών και σε εκχυλίσματα κυττάρων χωρίς κύτταρα HeLa αλλά όχι σε κυτταρόλυμα δικτυοκυττάρων κουνελιού. J Virol 85: 638-643. doi: 10.1128 / JVI.01806-10
126.
Καμιτάνη W, Huang C, Narayanan K et al (2009) Μια στρατηγική δύο επιπέδων για την καταστολή της πρωτεϊνικής σύνθεσης του ξενιστή από την πρωτεΐνη Nsp1 του coronavirus SARS. Nat Struct Μοί ΒίοΙ 16: 1134-1140. doi: 10.1038 / nsmb.1680
127.
Ο Kamitani W, Narayanan Κ, Huang C et al (2006). Η σοβαρή οξεία αναπνευστική νόσο του κοροναϊού nsp1 καταστέλλει την έκφραση γονιδίου ξενιστή προάγοντας την αποικοδόμηση του mRNA του ξενιστή. Proc Natl Acad Sci USA 103: 12.885 - 12.890
128.
(2012) Το σοβαρό οξύ αναπνευστικό σύνδρομο του κοροναϊού nsp1 διευκολύνει την αποτελεσματική διάδοση στα κύτταρα μέσω ενός ειδικού μεταφραστικού τερματισμού του mRNA του ξενιστή. J Virol 86: 11128-11137. doi: 10.1128 / JVI.01700-12
129.
Graham RL, Sims AC, Brockway SM et al (2005) Οι πρωτεΐνες replicase nsp2 του ιού της ηπατίτιδας μυοειδούς και του κοροναϊού σοβαρής οξείας αναπνευστικής σύνθεσης διατίθενται για ιική αντιγραφή. J Virol 79: 13399-13411. doi: 10.1128 / JVI.79.21.13399-13411.2005
130.
Η συνθετική πρωτεΐνη 2 σοβαρής αναπνευστικής λειτουργίας του κοροναϊού, αλληλεπιδρά με ένα σύμπλεγμα ξενιστικών πρωτεϊνών που εμπλέκεται στη βιογενέση των μιτοχονδρίων και στην ενδοκυτταρική σηματοδότηση. J Virol 83: 10314-10318. doi: 10.1128 / JVI.00842-09
131.
Η δομή πυρηνικού μαγνητικού συντονισμού δείχνει ότι η περιοχή του σοβαρού οξέως αναπνευστικού συνδρόμου του κορωναϊού περιέχει μια πτυχή macrodomain. J Virol 83: 1823-1836
132.
Egloff MP, Malet Η, Putics Α et al (2006) Δομική και λειτουργική βάση για δέσμευση ADP-ριβόζης και πολυ (ADP-ριβόζης) από ιικά μακροσώματα. J Virol 80: 8493-8502. doi: 10.1128 / JVI.00713-06
133.
Η παθολογία του ήπατος του ιού της ηπατίτιδας του ποντικού εξαρτάται από την ADP-ριβόζη-1 "-φωσφατάση, μια ιϊκή λειτουργία διατηρούμενη στην άλφα-όμοια υποομάδα. J Virol 82: 12325-12334. doi: 10.1128 / JVI.02082-08
134.
(2009) Το σοβαρό οξύ αναπνευστικό σύνδρομο coronavirus-ομοιάζον με παπαϊνη πρωτεάση περιοχή που μοιάζει με ουβικιτίνη και ο καταλυτικός τομέας ρυθμίζουν τον ανταγωνισμό της σηματοδότησης IRF3 και NF-kappaB. J Virol 83: 6689-6705
135.
Neuman BW, Joseph JS, Saikatendu KS et al (2008) ανάλυση Πρωτεομική ξετυλίγεται το λειτουργικό ρεπερτόριο του κορωνοϊού μη δομικής πρωτεΐνης 3. J Virol 82: 5.279-5294
136.
Serrano P, Johnson ΜΑ, Almeida MS et al (2007) Δομή πυρηνικού μαγνητικού συντονισμού του Ν-τερματικού πεδίου της μη δομικής πρωτεΐνης 3 από το σοβαρό οξείδιο του αναπνευστικού συνδρόμου κοροναϊό. J Virol 81: 12049-12060
137.
Serrano P, Johnson ΜΑ, Chatterjee Α et al (2009) Δομή πυρηνικού μαγνητικού συντονισμού της περιοχής δέσμευσης νουκλεϊκού οξέος της μη δομικής πρωτεΐνης σοβαρής οξείας αναπνευστικού συνδρόμου coronavirus 3. J Virol 83: 12998-13008. doi: 10.1128 / JVI.01253-09
138.
Ziebuhr J, Thiel V, Gorbalenya ΑΕ (2001) Η αυτοκαταλυτική απελευθέρωση ενός υποτιθέμενου παράγοντα μεταγραφής του ιού RNA από τον πολυπρωτεϊνικό πρόδρομο περιλαμβάνει δύο παράλογες πρωτεάσες τύπου παπαϊνης οι οποίες διασπούν τον ίδιο πεπτιδικό δεσμό. J Biol Chem 276: 33220-33232. doi: 10.1074 / jbc.M104097200
139.
Ο Clementz MA, Kanjanahaluethai A, O'Brien TE et al (2008) Η μετάλλαξη στην πρωτεΐνη nsp4 αντιγραφής του κορδονειδούς ποντικού μεταβάλλει τη συγκρότηση κυστιδίων διπλής μεμβράνης. Ιολογία 375: 118-129
140.
Gradge MJ, Sparks JS, Beachboard DC et al (2010) Η μη-δομική πρωτεΐνη του ιού της ηπατίτιδας ποντικού 4 ρυθμίζει τις τροποποιήσεις της μεμβράνης που προκαλούνται από τον ιό και τη σύνθετη λειτουργία του αναδιπλασιασμού. J Virol 84: 280-290. doi: 10.1128 / JVI.01772-09
141.
Lu Y, Lu X, Denison MR (1995) Ταυτοποίηση και χαρακτηρισμός μιας πρωτεϊνάσης τύπου σερίνης του μυϊκού κορωναϊού MHV-A59. J Virol 69: 3554-3559
142.
Oostra M, Hagemeijer MC, van Gent M et al (2008) Τοπολογία και μεμβρανική αγκύρωση του συγκροτήματος αντιγραφής του κοροναϊού: δεν είναι όλες οι υδρόφοβες περιοχές των nsp3 και nsp6 που καλύπτουν μεμβράνες. J Virol 82: 12392 με 12405
143.
Zhai Y, Sun F, Li X et al (2005) Αναλύει την μεταγραφή και αναπαραγωγή SARS-CoV από τη δομή του hexadecamer nsp7-nsp8. Nat Struct Μοί ΒίοΙ 12: 980-986
144.
Imbert I, Guillemot JC, Bourhis JM et al (2006) Μια δεύτερη μη-κανονική RNA-εξαρτώμενη RNA πολυμεράση σε SARS coronavirus. ΕΜΒΟ J 25: 4933-4942
145.
(2004) Η αντιγραφή πρωτεΐνης nsp9 της σοβαρής οξείας αναπνευστικού συνδρόμου-κοροναϊού είναι μία μονόκλωνη υπομονάδα δέσμευσης RNA μοναδική στον κόσμο του ιού RNA. Proc Natl Acad Sci USA 101: 3792 - 3796
146.
Bouvet M, Debarnot C, Imbert Ι et al (2010) In vitro ανασύσταση της μεθυλίωσης του πώλου mRNA του SARS-coronavirus. PLoS Pathog 6: e1000863. doi: 10.1371 / journal.ppat.1000863
147.
(2011) Κρυσταλλική δομή και λειτουργική ανάλυση του σύμπλοκου 2'-Ο-μεθυλοτρανσφεράσης nsp10 / nsp16 του καψιδίου SARS-κοροναϊού. PLoS Pathog 7: e1002059. doi: 10.1371 / journal.ppat.1002059
148.
Xu X, Liu Y, Weiss S et al (2003) Μοριακό μοντέλο πολυμεράσης κορωνοαϊού SARS: επιπτώσεις για βιοχημικές λειτουργίες και σχεδιασμό φαρμάκων. Νουκλεϊνικών οξέων Res 31: 7117 έως 7130
149.
Ivanov KA, Thiel V, Dobbe JC et al (2004) Πολλαπλές ενζυματικές δραστηριότητες που σχετίζονται με σοβαρή οξεία αναπνευστική σύνδρομο ελικάση κοροναϊού. J Virol 78: 5619-5632
150.
Ivanov KA, Ziebuhr J (2004) Ανθρώπινο κοροναϊό 229Ε μη δομική πρωτεΐνη 13: χαρακτηρισμός των δραστικοτήτων διπλής εκτομής, νουκλεοζιτικής τριφωσφατάσης και 5'-τριφωσφατάσης RNA. J Virol 78: 7833-7838. doi: 10.1128 / JVI.78.14.7833-7838.2004
151.
Ο Eckerle LD, Becker MM, Halpin RA et al (2010) Η απιστία της αναπαραγωγής του ιού μεταλλαγμένου ιού του SARS-CoV Nsp14-εξωνουκλεάσης αποκαλύπτεται με πλήρη αλληλούχιση γονιδιώματος. PLoS Pathog 6: e1000896. doi: 10.1371 / journal.ppat.1000896
152.
Eckerle LD, Lu X, Sperry SM et al (2007) Η υψηλή πιστότητα της αντιγραφής του ιού της ηπατίτιδας των ποντικών μειώνεται σε μεταλλάξεις nsp14 εξωριβονουκλεάσης. J Virol 81: 12135 έως 12144
153.
Minskaia E, Hertzig Τ, Gorbalenya AE et al (2006) Ανακάλυψη ενός εξωριβονουκλεάσης 3 '-> 5' εξωριβονουκλεάσης RNA που εμπλέκεται κρίσιμα στη σύνθεση RNA κοροναϊού. Proc Natl Acad Sci USA 103: 5108-5113. doi: 10.1073 / pnas.0508200103
154.
Chen Y, Cai H, Pan J et al (2009) Η λειτουργική οθόνη αποκαλύπτει τη μη δομική πρωτεΐνη nsp14 του κοραλοειδούς του SARS ως νέα κεφαλή N7 μεθυλοτρανσφεράση. Proc Natl Acad Sci USA 106: 3484-3489. doi: 10.1073 / pnas.0808790106
155.
Bhardwaj K, Sun J, Holzenburg Α et al (2006) RNA αναγνώριση και διάσπαση από την endoribonuclease του coronavirus SARS. J ΜοΙ ΒίοΙ 361: 243-256. doi: 10.1016 / j.jmb.2006.06.021
156.
Οι Ivanov KA, Hertzig T, Rozanov M et al (2004) Κύριος γενετικός δείκτης νινοϊών κωδικοποιεί μια αντιγραφόμενη endoribonuclease. Proc Natl Acad Sci USA 101: 12694-12699
157.
Οι Decroly E, Imbert Ι, Coutard Β et αϊ (2008) Η μη δομική πρωτεΐνη 16 του κορωναϊού είναι ένα ένζυμο πρόσδεσης cap-0 που έχει δραστικότητα (νουκλεοσιδίου-2'Ο) -μεθυλοτρανσφεράσης. J Virol 82: 8071 έως 8084
158.
(2) Ο-μεθυλίωση της ριβόζης παρέχει μια μοριακή υπογραφή για τη διάκριση του εαυτού και του μη-mRNA που εξαρτάται από τον αισθητήρα RNA Mda5. Nat Immunol 12: 137-143


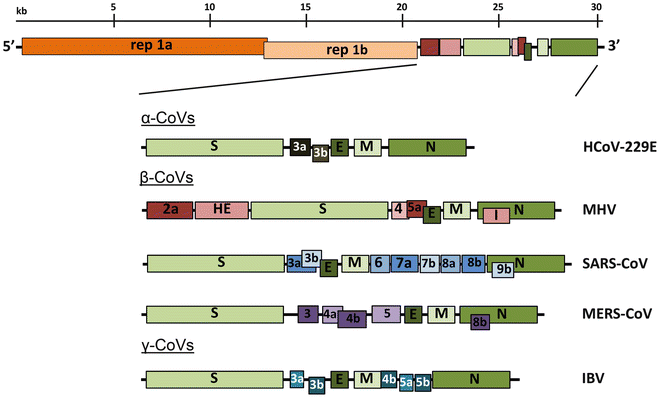
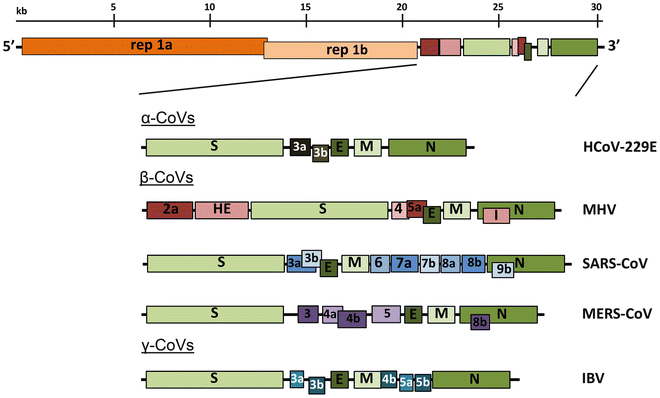
Δεν υπάρχουν σχόλια:
Δημοσίευση σχολίου